Mamíferos mariños
Os mamíferos mariños son mamíferos acuáticos que dependen dos ecosistemas mariños para a súa existencia. Son os pinnípedes (focas, leóns mariños, morsas), cetáceos (baleas, golfiños, toniñas), sirenios (manatís, dugongos), londras mariñas e osos polares. Non representan un taxon diferenciado ou agrupación sistemática, senón que teñen unha relación polifilética debida a evolución converxente, dado o seu tipo de vida similar, pero non teñen un antepasado común próximo.

A adaptación dos animais mariños a un estilo de vida acuático varía considerablemente entre as especies. Tanto os cetáceos coma os sirenios son completamente acuáticos e nunca visitan a terra firme. As focas e leóns mariños son semiacuáticos, xa que, aínda que pasan a maior parte do seu tempo na auga, tamén necesitan volver a terra para reproducirse, descansar, mudar. Ao contrario dos anteriores, as londras mariñas e os osos polares están moito menos adaptados á vida acuática. As dietas dos mamíferos mariños varían tamén considerablemente; algúns comen zooplancto, outros comen peixes, luras, moluscos, plantas mariñas e uns poucos comen outros mamíferos. Aínda que o número de mamíferos mariños é pequeno comparado cos que hai en terra, a súa función en diversos ecosistemas é importante, especialmente no que concirne ao mantemento dos ecosistemas mariños, por exemplo, ao regularen as poboacións das súas presas. Este papel no mantemento dos ecosistemas fai que cause preocupación que un 23% das especies mariñas estean ameazadas actualmente.

Os mamíferos mariños foron cazados desde hai moito tempo polos pobos aborixes para obter alimento e outros recursos. Moitos foron tamén o obxectivo da industria comercial, o que orixinou un acusado declive en todas as poboacións das especies explotadas, como as baleas e focas. A caza comercial levou á extinción da Hydrodamalis gigas e da Neomonachus tropicalis. Unha vez que cesou a caza comercial, algunhas especies como a balea cincenta e o elefante mariño do norte, volveron a aumentar en número; inversamente, outras especies, como a balea vasca, están en perigo crítico. Ademais de por causa da caza, os mamíferos mariños poden morrer como capturas accidentais (bycatch) en bancos de pesca, ao quedaren enguedellados nas redes de enmalle e afogaren alí ou morreren de fame. O incremento do tráfico marítimo causa con frecuencia colisións entre os buques oceánicos rápidos e os grandes mamíferos mariños. A degradación do hábitat tamén ameaza os mamíferos mariños e a súa capacidade de atopar e capturar alimentos. A contaminación acústica, por exemplo, pode afectar adversamente á ecolocalización que usan os mamíferos, e os crecentes efectos do quecemento global degradan o medio ambiente ártico, onde moitos deles viven.
Taxonomía editar




.jpg)
Clasificación das especies existentes editar
| Filoxenia dos mamíferos mariños | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Os taxons en grosa son mariños.[1] |
- Orde Cetartiodactyla[2]
- Suborde Whippomorpha
- Familia Balaenidae (baleas), dous xéneros e catro especies
- Familia Cetotheriidae (balea anana), unha especie
- Familia Balaenopteridae (rorcuais), dous xéneros e oito especies
- Familia Eschrichtiidae (balea cincenta), unha especie
- Familia Physeteridae (cachalote), unha especie
- Familia Kogiidae (cachalotes pigmeos e ananos), un xénero e dúas especies
- Familia Monodontidae (narval e beluga), dous xéneros e dúas especies
- Familia Ziphiidae (zífidos), seis xéneros e 21 especies
- Familia Delphinidae (golfiños mariños), 17 xéneros e 38 especies
- Familia Phocoenidae (toniñas), dous xéneros e sete especies
- Suborde Whippomorpha
- Orde Sirenia [2]
- Suborde Cynodontia
- Familia Trichechidae (manatís), un xénero e tres especies
- Familia Dugongidae (dugongos), unha especie
- Suborde Cynodontia
- Orde Carnivora [2]
- Suborde Caniformia
- Familia Mustelidae, (londras mariñas norteamericana e suramericana), dous xéneros e dúas especies
- Familia Ursidae (oso polar), unha especie
- Suborde Pinnipedia
- Familia Otariidae (leóns mariños), 7 xéneros e 15 especies
- Familia Odobenidae (morsas), unha especie
- Familia Phocidae (focas), 14 xéneros e 18 especies
- Suborde Caniformia
Evolución editar

Os mamíferos mariños forman un grupo diverso de 129 especies que dependen do océano para a súa alimentación e existencia.[3][4] Non representan un taxon específico ou agrupación sistemática, senón que teñen relacións polifiléticas.[5] Malia a súa diversidade de morfoloxías entre os distintos grupos, a mellora da súa eficiencia na busca de alimentos foi o principal impulsor na súa evolución.[6][7] O seu nivel de dependencia do medio mariño para vivir varía considerablemente coa especie. Os golfiños e baleas son completamente dependentes do medio ambiente mariño en todas as etapas da súa vida. As focas aliméntanse no océano pero reprodúcense en terra. Os osos polares deben alimentarse en terra.[5] O 23% das especies de mamíferos mariños son especies ameazadas.[8][9]
Os cetáceos fixéronse acuáticos hai uns 50 millóns de anos.[10] Baseándose en investigacións moleculares e morfolóxicas, os cetáceos están xenética e morfoloxicamente dentro do grupo dos Artiodactyla (ungulados de dedos pares).[11][12] O termo "Cetartiodactyla" indica a idea de que as baleas evolucionaron dentro dos ungulados. O termo foi acuñado ao fusionar o nome das dúas ordes, Cetacea e Artiodactyla, nunha soa palabra. Con esta definciión, o parente terrestre máis próximo vivo das baleas e golfiños crese que é o hipopótamo.[13][14][15][16]
Os sirenios fixéronse acuáticos hai uns 40 millóns de anos. A primeira aparición dos sirenios no rexistro fósil foi durante o Eoceno temperán, e no Eoceno tardío os sirenios xa estaban bastante diversificados. Son habitantes de ríos, estuarios e augas mariñas costeiras, e poden espallarse rapidamente. Os restos do sirenio máis primitivo do que se ten noticia, †Prorastomus, atopáronse en Xamaica,[7] a diferenza doutros mamíferos mariños que se orixinaron no Vello Mundo (como os cetáceos[17]). O primeiro sirenio cuadrúpede coñecido foi †Pezosiren do Eoceno temperán.[18] As familias máis antigas coñecidas de sirenios, †Prorastomidae e †Protosirenidae, estaban confinadas no Eoceno, eran do tamaño dun porco, tiñan catro patas, e eran criaturas anfibias.[19] Os primeiros membros dos Dugongidae apareceron ao final do Eoceno.[20] Nese momento, xa eran completamente acuáticas.[19]
Os pinnípedes separáronse doutros caniformes hai 50 millóns de anos durante o Eoceno. A súa ligazón evolutiva con mamíferos terestres era descoñecida ata que se descubriu en 2007 †Puijila darwini en depósitos do Mioceno temperán de Nunavut, Canadá. Igual que hoxe ocorre coas londras modernas, aqueles antepasados dos pinnípedes do xénero †Puijila tiñan colas longas, patas curtas e pés palmados en vez de aletas.[21] As liñaxes dos Otariidae (leóns mariños) e Odobenidae (morsas) separáronse hai case 28 millóns de anos.[22] Os fócidos (focas) levan existindo desde hai polo menos 15 millóns de anos[23] e as probas moleculares apoian a diverxencia das liñaxes dos Monachinae (focas monxe) e dos Phocinae hai 22 millóns de anos.[22]
As evidencias fósiles indican que a liñaxe da londra mariña (Enhydra) quedou illada no norte do Pacífico hai aproximadamente dous millóns de anos, dando lugar ao agora extinto †Enhydra macrodonta e a moderna londra mariña, Enhydra lutris. A londra mariña evolucionou inicialmente no norte de Hokkaidō e Rusia, e despois espallouse cara ao leste ás illas Aleutianas, as costas continentais de Alasca, e cara ao sur pola costa de Norteamérica. En comparación cos cetáceos, sirenions e pinnípedes, que entraron na auga aproximadamente 50, 40, e 20 millóns de anos, respectivamente, a londra de mar é relativamente unha acabada de chegar á vida mariña. Porén, en certos aspectos, a londra mariña está máis completamente adaptada á auga que os pinnípedes, que deben arrastrarse a terra ou sobre o xeo para dar a luz.[24]
Os osos polares pénsase que diverxiron dunha poboación de osos pardos da especie Ursus arctos, que quedaron illados durante un período de glaciación no Plistoceno[25] na parte leste de Siberia, (desde Kamchatka e a península de Kolym).[26] O fósil de oso polar máis antigo coñecido é unha mandíbula de hai de 130 000 a 110 000, atopada na illa Prins Karls Forland (de Svalbard) en 2004.[27] O ADN mitocondrial do oso polar diverxiu do oso pardo hai uns 150 000 anos.[27] Ademais, algúns clados de oso pardo, como se estima polos seus ADN mitocondriais, están máis estreitamente relacionados cos osos polares que con outros osos pardos,[28] o que significa que o oso polar non debería considerarse unha especie se se seguen certas definicións de especie.[29]
Distribución e hábitat editar

Os mamíferos mariños están distribuídos por todo o globo, pero a súa distribución é irregular e coincide coa produtividade nos océanos.[31] A riqueza de especies é máxima arredor dos 40° de latitude, tanto norte coma sur. Isto corresponde cos maiores niveis de produción primaria arredor de Norteamérica e Suramérica, África, Asia e Australia. A área de distribución total das especies é moi variable. Como media a maioría dos mamíferos mariños ten áreas de distribución que son equivalentes ou menores que da quinta parte do océano Índico.[8] A variación observada no tamaño da área de distribución é o resultado das diferentes necesidades ecolóxicas de cada especie e das súas capacidades de enfrontarse a un amplo rango de condicións ambientais. Preocupa que exista un alto grao de solapamento entre a riqueza de especies de animais mariños e as áreas de impacto humano sobre o medio ambiente.[3]
A maioría dos mamíferos mariños, como focas e londras mariñas, habitan nas costas. Porén, as focas tamén utilizan diversos hábitats terrestres, tanto continentais coma en illas. En áreas tropicais e temperadas, arrástranse a praias de area ou croios, costas rochosas ou escollos, chairas limosas costeiras, pozas de marea e covas mariñas. Algunhas especies descansan en estruturas construídas polo ser humano, como peiraos, escolleiras, boias e plataformas petrolíferas. As focas poden internarse algo máis en terra e descansar en dunas ou vexetación, e poden incluso subir por cantís.[32]:96 A maioría dos cetáceos viven no océano aberto, e especies como os cachalotes poden mergullarse a profundidades de 300 a 760 m para procurar alimento.[33] Os sirenios viven en augas costeiras pouco fondas, xeralmente de 9,1 :m de profundidade. Porén, sábese que poden mergullarse a 37 m de profundidade para alimentarse das herbas mariñas que medran a maior profundidade.[34] As londras mariñas viven en áreas protexidas, como costas rochosas, bosques de kelp e arrecifes de coral,[35] aínda que poden vivir entre o xeo á deriva ou en áreas con limo, lama ou area.[36]
Moitos animais mariños migran estacionalmente. En moitas áreas as zonas de xeo aparecen e desaparecen anualmente cos cambios de temperatura, e as focas migran en resposta a eses cambios. Á súa vez, os osos polares deben seguir as súas presas nas súas migracións. Na baía de Hudson, baía de James e outras áreas, o xeo derrétese completamente cada verán forzando os osos polares a abandonar a zona onde estaban os xeos e ir a terra firme e esperar meses ata que se produza unha nova conxelación. No mar de Chukchi e no de Beaufort, os osos polares retíranse todos os veráns cara ao xeo que hai máis ao norte, que permanece conxelado todo o ano.[37] As focas poden tamén migrar por causa doutros cambios ambientais, como o fenómeno climático de El Niño, e as focas viaxeiras poden utilizar varias características das súas contornas para conseguir chegar aos seus destinos, como os campos xeomagnéticos, as correntes de auga e os ventos, a posición do Sol e a Lúa e o sabor e temperatura da auga.[32]:256–257 As baleas misticetas fan grandes migracións cubrindo grandes distancias a augas tropicais para daren a luz e criaren os fillos,[38] posiblemente para impedir a predación polas candorcas.[39] A balea cincenta fai a migración máis longa rexistrada nun animal, viaxando 23 000 km desde o mar de Okhotsk á Península de Baixa California.[40] Durante o inverno, os manatís que viven no extremo norte da súa área de distribución migran a augas máis cálidas.[41]
Adaptacións editar
- Véxase tamén: Locomoción acuática.

Os mamíferos mariños teñen diversas características fisiolóxicas e anatómicas que lles permiten superar os retos asociados coa vida acuática. Algunhas destas características son moi específicas de especie. Desenvolveron tamén características especiais para unha locomoción eficiente, como formas corporais de torpedo para reducir a resistencia; membros modificados para a propulsión e guía; aletas caudais e dorsais para a propulsión e o equilibrio.[31] Os mamíferos mariños termorregúlanse usando un pelame denso ou capas de graxa, axustando a súa circulación (intercambio de calor contracorrente); corpos con forma de torpedo, apéndices reducidos, e un gran tamaño para reducir as perdas de calor.[31]
Os mamíferos mariños poden mergullarse durante longos períodos. Tanto pinnípedes coma cetáceos teñen sistemas de vasos sanguíneos grandes e complexos, que serven para almacenar oxíxeno para poder soportar mergullarse a gran profundidade. Outras importantes reservas de oxíxeno son os músculos, sangue e bazo. Tamén poden reducir moito a súa frecuencia cardíaca (bradicardia) e realizar unha vasoconstrición (que desvía a maioría do oxíxeno aos órganos vitais, como cerebro e corazón) para poder aumentar o tempo que pasan mergullados e soportar a privación de oxíxeno.[31] Se o oxíxeno se vai esgotando, os mamíferos mariños poden acceder a reservas substanciais de glicóxeno que alimentan a glicólise anaerobia das células, que se activa durante as condicións de hipoxia sistémica durante os mergullos prolongados.[42][43][44]
O son transmítese de forma diferente pola auga que polo aire, polo que os mamíferos mariños desenvolveron adaptacións para asegurar a comunicación efectiva, captura de presas e detección de predadores.[45] A adaptación máis salientable é o desenvolvemento da ecolocalización en baleas e golfiños.[31] Os odontocetos emiten un chorro enfocado de clics de alta frecuencia na dirección en que apunta a súa cabeza. Os sons xéranse facendo pasar aire polas narinas óseas a través dos labios fónicos.[46] Estes sons son reflectidos polo óso cóncavo denso do cranio e un saco de aire na súa base. O chorro enfocado de clics é modulado por un órgano graxo grande chamado 'melón'. Este actúa como unha lente acústica porque está composto de lípidos de diferentes densidades.[47]
Nos mamíferos mariños evolucionaron moitas características para mellorar a súa alimentación, como se poden ver na súa dentición. Por exemplo, os dentes da zona da meixela dos pinnípedes e odontocetos están especialmente adaptados para a captura de peixes e luras. En contraste, os misticetos teñen barbas de balea para filtrar plancto e pequenos peixes.[31]
Os osos polares, londras mariñas e focas teñen mestos pelames, con capas de pelos longos, aceitosos e impermeables para atrapar o aire e proporcionar illamento. En contraste, outros mamíferos mariños, como as baleas, golfiños, toniñas, manatís, dugongos e morsas, perderon os pelos e no seu lugar teñen unha epiderme especialmente grosa e densa e unha capa de graxa máis grosa baixo a pel en resposta ás súas necesidades hidrodinámicas. Os animais que se alimentan vadeando e nos fondos (como os manatís) necesitan ser máis pesados que a auga para manterse en contacto co fondo ou permanecer mergullados. Os animais que viven na superficie (como as londras mariñas) necesitan o contrario, e os animais que nadan libremente en augas abertas (como os golfiños) necesitan ter unha flotabilidade neutra para poder nadar arriba e abaixo na columna de auga. Tipicamente, os ósos densos e grosos encóntranse en animais que se alimentan no fondo, mentres que unha densidade ósea baixa está asociada con mamíferos que non viven en augas profundas. Algúns mamíferos mariños, como os osos polares e londras, conservan patas fortes que sosteñen o seu peso e poden camiñar por terra como os animais completamente terrestres.[48]
Ecoloxía editar
Dieta editar

Todos os cetáceos son carnívoros e predadores. Os odontocetos aliméntanse principalmente de peixes e cefalópodos, seguidos de crustáceos e bivalvos. Algúns poden comer outro tipo de animais, como outras especies de baleas ou certas especies de pinnípedes.[32]:169[49] Un método común de alimentación é cazar en mandas, na cal un grupo de golfiños acosa e "comprime" un banco de peixes para que teña un menor volume, formando o que se coñece como bóla de isco. Cada membro da manda fai quendas arremetendo contra a bóla, e comendo os peixes que quedan atordados.[50] Outro método usado polos golfiños é acurralar os peixes en augas pouco profundas, onde os poden capturar máis doadamente.[50] As candorcas e golfiños Tursiops tamén dirixen as presas cara a praias par alimentarse delas.[51][52] Outras baleas de fociño romo e dentición reducida aliméntanse por succión.[53] Aínda que son animais carnívoros, albergan unha flora intestinal similar á dos herbívoros terrestres, probablemente un legado dos seus antepasados herbívoros.[54]
Os misticetos usan as súas placas de barbas de balea para filtrar da auga plancto e outros pequenos animais; usan dous tipos e métodos: alimentarse con arremetidas ou tragando auga. Os que se alimentan por arremetidas abren as súas mandíbulas e expanden boca ata un volume moito maior do orixinal, xa que as engurras da súa gorxa se estiran, aumentando moito a cantidade de auga que poden albergar na boca.[55][56] Turran contra unha bóla de isco a gran velocidade, pero isto só é efectivo enerxeticamente cando se trata dunha bóla grande. Despois filtran a auga coas barbas para que quede na boca o alimento e non tragar toda a auga.[57] Os que se alimentan tragando auga nadan coa boca aberta, que se vai enchendo de presas e auga. As presas deben aparecer en gran cantidade para que teñan interese para a balea, estar dentro de certo tamaño para que as barbas da balea a poidan filtrar, e ser o suficientemente lentas para que non lles dea tempo a escapar.[58] Algunhas especies aliméntanse en grupo facendo unha parede circular de burbullas para concentrar as presas no centro e ascendendo polo centro coa boca aberta.

As londras mariñas son os únicos animais mariños que poden levantar e dar volta a pedras, o que fan coas patas anteriores cando están buscando presas polo fondo do mar.[59] Poden apañar os caracois e outros organismos mariños pegados ao kelp e escavar no barro do fondo do mar para buscar ameixas.[59] É o único animal mariño que captura peixes coas poutas das patas anteriores en vez de usar os dentes.[60] Baixo cada unha das patas anteriores, as londras de mar teñen unha bolsa frouxa de pel que se estende ata o peito, que usan para almacenar o alimento que recolleron e levalo á superficie. Nesta bolsa tamén poden gardar unha pedra que se utiliza para romper as cunchas de ameixas, o que é un exemplo de uso de ferramentas por un animal.[61] As londras de mar comen mentres aboian boca arriba, usando as poutas das patas anteriores para desgarrar a comida e levala a boca.[62][63] As londras mariñas aliméntanse principalmente de crustáceos e peixes.[64]
Os pinnípedes aliméntanse principalmente de peixes e cefalópodos, en segundo lugar de crustáceos e bivalvos e despois de zooplancto e nalgúns casos de presas de sangue quente (como aves mariñas).[32]:145 A maioría das especies son xeralistas na súa alimentación, pero hai unhas poucas especialistas.[65] Normalmente cazan peixes que non forman bancos, invertebrados inmóbiles ou lentos ou presas endotérmicas cando están en grupos. As especies que se alimentan en solitario xeralmente fano nas augas costeiras, baías e ríos. Cando están dispoñibles grandes bancos de peixes ou luras, os pinnípedes cazan cooperativamente en grandes grupos, localizando e agrupando as súas presas. Algunhas especies, como os leóns mariños de California e de Suramérica, poden alimentarse de cetáceos e aves mariñas.[32]:168
O oso polar é a especie máis carnívora de óso, e a súa dieta consiste principalmente en focas das especies Pusa hispida e Erignathus barbatus.[66] Os osos polares cazan principalmente na interface entre o xeo e a auga e aire; raramente cazan focas en terra e moito menos en augas abertas.[67] O método de caza máis común dos osos polares é a caza silandeira:[68] O oso localiza o burato respiratorio no xeo dunha foca usando a seu sentido do olfacto, e espera preto a que a foca apareza. Cando a foca exhala, o oso chéiraa e cóllea no burato coas garras das patas anteriores, e arrástraa ao xeo. O oso polar tamén caza sorprendendo as focas que descansan no xeo. Unha vez que enxergan unha foca, camiñan achegándose a uns 90 m, e agóchase contra o chan. Se as focas non se decatan da súa presenza, o oso arrástrase ata uns 9 ou 10 m da foca e despois lanza de repente un rápido ataque.[69] Un terceiro método de caza é asaltar as toqueiras de cría que as focas femias escavaron na neve.[68] Poden tamén alimentarse de peixes.[70]

Os sirenios denomínase ás veces "vacas mariñas" porque a súa dieta consiste principalmente en herbas mariñas. Cando comen, inxiren a planta completa, incluíndo as raíces, aínda que cando isto lles é imposible, comen só as follas.[71] Nos contidos estomacais dos dugongos atopáronse unha ampla variedade de herbas mariñas (anxiospermas), e existen probas de que comen tamén algas cando as herbas mariñas escasean.[72] O manatí Trichechus manatus come unhas 60 especies de plantas, e en menor medida tamén peixes e pequenos invertebrados.[73]
Especies clave editar
- Véxase tamén: Especie chave.
As londras mariñas son un exemplo clásico de especie clave; a súa presenza afecta o ecosistema máis profundamente que o que indicarían os seus tamaños e cantidades. Manteñen controladas as poboacións de certos herbívoros bénticos, especialmente os ourizos de mar. Os ourizos de mar pastan os talos inferiores do kelp, causando que os kelp queden á deriva e morran. A perda de hábitats e nutrientes que provían os bosques de kelp orixinan uns profundos efectos fervenza no ecosistema mariño. En áreas do norte do Pacífico onde non hai londras mariñas toda a zona se converteu nun "ermo de ourizos", cunha grande abundancia de ourizos de mar, pouca biodiversidade e ningún bosque de kelp.[74] A reintrodución de londras de mar na Columbia Británica causou unha drástica mellora na saúde dos ecosistemas costeiros,[75] e observáronse cambios similares a medida que as poboacións de londras mariñas se recuperaron nas illas Aleutianas e Commander e a costa do Gran Sur de California.[63] Porén, algúns ecosistemas de bosque de kelp de California tamén conseguiron prosperar sen a presenza de londras mariñas, xa que as poboacións de ourizos foron controladas aparentemente por outros factores.[63] O papel das londras de mar para manter os bosques de kelp é máis importante en áreas de costa aberta que en baías abrigadas e estuarios.[63]
.jpg)

Un superdepredador afecta á dinámica das poboacións de presas e as súas tácticas de defensa (como o camuflaxe).[76] O oso polar é un superdepredador no seu ámbito.[77] Varias especies animais, especialmente o raposo ártico (Vulpes lagopus) e a gaivota Larus hyperboreus, comen rutineiramente prea das presas que o oso polar matou.[78] As relacións entre as focas Pusa hispida e os osos polares é tan próxima que a abundancia desas focas nalgunhas áreas parece regular a densidade de osos polares, mentres que a predación dos osos á súa vez regula a densidade e éxito reprodutor das focas.[79] A presión evolutiva da predación dos osos polares sobre as focas probablemente explica algunhas diferenzas significativas entre as focas antárticas e árticas. Comparadas coas antárticas, onde non hai un depredador importante de superficie, as focas árticas usan máis buratos respiratorios no xeo por individuo, parecen máis móbiles cando se arrastran sobre o xeo e raramente defecan no xeo.[78] O pelame das crías das focas árticas é branca, presumiblemente para proporcionarlles camuflaxe contra os depredadores, mentres que as crías antárticas teñen todas pelo escuro.[78]
As candorcas son tamén superdepredadores con distribución global, e poden ter un profundo efecto sobre o comportamento e as poboacións das especies de presas. As súas dietas son moi amplas e poden alimentarse con moitos vertebrados do océano incluíndo o salmón,[80] raias, tiburóns (incluído o tiburón branco),[81][82] grandes misticetos,[83] e case 20 especies de pinnípedes.[84] A predación de crías de balea pode ser responsable das migracións anuais das baleas a zonas de cría en augas máis tropicais, onde a poboación de candorcas é moito menor que nas augas polares. Antes da caza de baleas, crese que as grandes baleas eran unha importante fonte de alimento para elas, porén, despois do seu forte declive, as condorcas ampliaron a súa dieta, o que orixinou unha diminución da abundancia de animais mariños máis pequenos.[39] Un declive de londras mariñas nas illas Aleutianas na década de 1990 foi controvertidamente atribuída por algúns científicos á predación polas condorcas, aínda que non hai probas directas. Ao declive das londras mariñas seguiu un declive das poboacións das focas Phoca vitulina e do león mariño Eumetopias jubatus, as presas preferidas das candorcas, que puideron ser os substitutos das súas presas orixinais, agora reducidas pola caza industrial das baleas.[85][86][87]
Reciclaxe de nutrientes editar
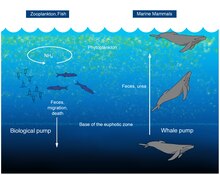
Un estudo de 2010 considerou que as baleas tiñan unha positiva influencia na produtividade das pesquerías oceánicas, xa que funcionaban como unha bomba de nutrientes, ao que se lle deu o nome de "bomba das baleas". As baleas transportan os nutrientes como o nitróxeno desde as profundidades á superficie. Isto funciona como unha bomba biolóxica ascendente, o que inverte unha opinión anterior que consideraba que as baleas aceleraban a perda de nutrientes cara ao fondo. Esta entrada de nitróxeno no Golfo de Maine é maior que a entrada debida a todos os ríos combinados que desembocan nese golfo, unhas 23 000 toneladas ao ano.[88] As baleas defecan na superficie do océano; os seus excrementos son importantes para as pesquerías porque son ricos en nitróxeno e ferro. As feces de balea son líquidas e en vez de afundirse, permanecen na superficie, onde o fitoplancto as utiliza.[88][89]
Despois da súa morte, as carcasas de balea acaban por caer ao fondo dos océanos e proporcionan un hábitat para certas criaturas mariñas. As evidencias destes afundimentos de carcasas non nosos días e no rexistro fósil mostran que estas carcasas sustentan unha rica ensamblaxe de seres vivos, cunha diversidade global de 407 especies, comparable á doutros puntos quentes de biodiversidade neríticos, como as emanacións frías e fontes hidrotermais oceánicas.[90] A deterioración das carcasas de balea ocorre en tres etapas. Inicialmente, os organismos móbiles, como tiburóns e mixíns, aliméntanse dos tecidos brandos con grande rapidez nun período de varios meses a dous anos. Isto vai seguido da colonización de ósos e os sedimentos que os rodean (que conteñen tamén materia orgánica) por oportunistas do enriquecemento, como crustáceos e vermes poliquetos, durante un período de anos. Finalmente, as bacterias sulfofílicas reducen os ósos liberando sulfuro de hidróxeno permitindo o crecemento de organismos quimioautótrofos, que á súa vez, sosteñen outros organismos como mexillóns, ameixas, lapas, e caracois mariños. Esta etapa pode durar décadas e soster unha rica ensamblaxe de especies, como media 185 especies por sitio.[91]
Interaccións cos humanos editar
Ameazas editar
Explotación editar
- Véxase tamén: Caza de baleas e Caza de focas.

Os animais mariños foron cazados historicamente polos pobos aborixes que vivían nas costas para obter alimento e outros recursos. Esta caza de subsistencia aínda ocorre no Canadá, Groenlandia, Indonesia, Rusia, os Estados Unidos, e varias nacións do Caribe. Os efectos desta caza tan só son localizados, xa que se realiza a unha escala relativamente pequena.[31] Ao contrario, a caza comercial realizouse a moita maior escala e os mamíferos mariños foron enormemente explotados. Isto levou á extinción de Hydrodamalis gigas (ao que tamén contribuíu a caza de subsistencia) e da Neomonachus tropicalis.[31] Hoxe, as poboacións das especies que foron cazadas historicamente, como a Balaenoptera musculus (balea azul) e a Eubalaena japonica, son moito menores do que eran antes da caza comercial.[92] Como as baleas xeralmente teñen taxas de crecemento lentas, tardan moito en acadar a madurez sexual, e teñen un baixo rendemento reprodutor, a recuperción das poboacións foi moi lenta.[45]
Varias especies de baleas aínda son cazadas, a pesar da moratoria da prohibición de 1986 da caza de baleas segundo os termos da Comisión Baleeira Internacional (IWC). Só quedan dúas nacións que cacen comercialmente baleas: Noruega, onde se cazan varios centos de Balaenoptera acutorostrata cada ano; e Islandia, onde se estableceron cotas de 150 Balaenoptera physalus e 100 B. acutorostrata ao ano.[93][94] A caza de baleas tamén se segue realizando no Xapón, onde se cazan varios centos de baleas antárticas e do Pacífico norte ao ano, segundo eles afirman para a investigación científica de acordo coa moratoria.[92] Porén, o comercio ilegal de baleas e carne de golfiño é un mercado significativo no Xapón e algúns países.[95]

As pelaxes máis valiosas para ocomercio foron as das londras mariñas, especialmente a que habitaba nas costas de Norteamérica desde o río Columbia no sur á enseada de Cook no norte. A pelaxe da londra mariña californiana era menos aprezada e menos rendible. Despois de que a londra mariña do norte fose cazada ata a súa extinción local, os comerciantes de peles marítimos centráronse na londra californiana ata que esta tamén chegou case á extinción.[96] Os comerciantes marítimos de peles británicos e americanos levaban as súas peles ao porto chinés de Guangzhou (Cantón), onde operaban baixo o chamado Sistema cantonés. As peles da América rusa vendíanse principalmente a China a través da cidade comercial mongol de Kyakhta, que fora aberta ao comercio con Rusia polo Tatado de Kyakhta de 1727.[97]
A caza comercial de focas e leóns mariños foi historicamente tan importante como a industria baleeira. As especies explotadas incluían Pagophilus groenlandicus, Cystophora cristata), Pusa caspica, Mirounga, morsas e as nove especies de otáridos de pelame valioso (dos xéneros Arctocephalus e Callorhinus).[98] A escala da caza de focas diminuíu substancialmente despois da década de 1960,[99] despois de que o goberno canadense reducidse a duración da tempada de caza e implantase medidas para protexer as femias adultas.[100] Varias especies que eran explotadas comercialmente aumentaron as súas poboacións; por exemplo, os otáridos de pel valiosa antárticos pode que sexan hoxe tan numerosas como o eran antes da caza. Os elefantes mariños do norte (Mirounga angustirostris) foron cazados case ata a extinción a finais do século XIX, e só quedaron unhas poucas poboacións na illa de Guadalupe. Desde entón recolonizou boa parte da súa área de distribución histórica, pero a anterior redución da poboación orixinou un pescozo de botella xenético.[98] Inversamente, a foca monxe mediterránea foi extinguida na maior parte da súa anterior área, que se estendía desde o Mediterráneo ao mar Negro e o noroeste de África, e só queda no noroeste do Mediterráneo e partes do noroeste de África.[101]
Os osos polares son un trofeo de caza que se pode cazar no Canadá cun permiso especial e o acompañamento dun guía inuit. Este traballo de guías e venda de permisos pode ser un importante ingreso en pequenas comunidades locais, xa que se obtén máis por este traballo que por vender a pel do oso polar nos mercados. Os Estados Unidos, Rusia, Noruega, Groenlandia e o Canadá permiten a caza de subsistencia, e o Canadá dá permisos para conseguir trofeos de caza ás comunidades indíxenas. As peles de oso poden utilizarse con propósitos de subsistencia, como trofeos de caza, ou comercializarse en mercados.[102][103]
Tráfico marítimo e pesquerías editar

As capturas accidentais de mamíferos mariños nas pesquerías é frecuente. As redes de enmalle e de deriva causan a maior mortaldade en cetáceos e pinnípedes, porén, é tamén común que queden enredados en palangres e aparellos de arrastre e outros.[104] As redes de cerco para atúns son especialmente problemáticas para o enredamento de golfiños.[105] As capturas accidentais afectan a todos os cetáceos, pequenos e grandes, en todos os tipos de hábitats. Porén, os cetáceos máis pequenos e pinnípedos son máis vulnerables xa que o seu tamaño implica que é improbable que escapen unha vez quedaron atrapados e frecuentemente acaban afogando.[92] Aínda que os cetáceos máis grandes poden arrastrar as redes, estas ás veces permanecen firmemente enredadas no exemplar e poden impedir que o animal se alimente e ás veces chega a morrer de fame.[92] As redes abandonadas ou perdidas e palangres poden causar mortalidade por inxestión ou enredamento.[106] Os animais mariños tamén quedan enredados en redes de acuicultura, porén, son casos raros e non teñen impacto nas poboacións.[107]
Os choques contra barcos causan a morte de bastantes mamíferos mariños, especialmente de baleas.[92] En particular, os barcos comerciais rápidos como os portacontedores poden causar grandes danos ou a morte cando coliden cos mamíferos mariños. As colisións ocorren tanto con grandes buques comerciais coma con barcos de recreo e causan lesións a baleas ou outros cetáceos menores. A Eubalaena glacialis en perigo crítico está particularmente afectada polos choques contra barcos.[108] Os barcos dedicados ao turismo deseñados para a observación de baleas e golfiños poden tamén impactar negativamente nos mamíferos mariños ao interferiren cos seus comportamentos naturais.[109]
A industria das pesquerías non só ameaza os mamíferos mariños polas capturas accidentais, senón tamén pola competición pola comida. A explotación dos grandes bancos pesqueiros causou a diminución do stock de peixes que son importantes presas comidas polos mamíferos mariños. Os pinnípedes foron especialmente afectados pola perda directa das súas fontes de comida, o que nalgúns casos pode producir escaseza de alimentos e deficiencias na dieta,[110] fame entre os individuos novos e a redución do recrutamento dentro da poboación.[111] Cando os stocks de peixes diminúen, a competición entre os mamíferos mariños e as actividades pesqueiras orixinou nalgunhas ocasións conflitos. A práctica de sacrificar a grande escala poboacións de mamíferos mariños polas pesquerías comerciais iniciouse en varias áreas para protexer os stocks de peixes para o consumo humano.[112]
A acuicultura de mariscos aprópiase de espazos, o que orixina unha competencia polo espazo. Porén, hai pouca competición directa coa colleita de mariscos que fai a acuicultura.[107] Por outra parte, os mamíferos mariños capturan regularmente peixes das granxas de acuicultura, o cal crea problemas significativos para este tipo de acuicultores mariños. Aínda que xeralmente hai mecanismos legais deseñados para deter os mamíferos mariños, como as redes antipredadores ou aparellos que os acosan ou molestan, estes son con frecuencia matados ilegalmente.[107]
Perda e degradación de hábitats editar

A degradación do hábitat está causada esencialmente polas actividades humanas. Os animais mariños que viven nos ambientes costeiros son os que máis probablemente se ven afectados pola degradación e perda de hábitat. Os desenvolvementos como os emisarios submarinos de augas residuais, dispositivos ancorados no fondo, dragados, voaduras, verteduras, construción de portos, proxectos hidroeléctricos e de acuicultura degradan e ocupan hábitats valiosos.[45] Por exemplo, a acuicultura do marisco extensiva ocupa un espazo moi valioso utilizado polos animais costeiros para importantes actividades, como reproducirse, alimentarse ou descansar.[107]
Os contaminantes que se verten no medio mariño acumúlanse no corpo dos mamíferos mariños ao almacenárense nos seus tecidos adiposos.[45] Entre os contaminantes que se encontran nos tecidos dos mamíferos mariños están os metais pesados, como o mercurio e o chumbo, e tamén organoclorados e hidrocarburos aromáticos policíclicos.[45] Por exemplo, estes poden causar efectos perturbadores sobre o sistema endócrino;[106] alterar o sistema reprodutor, e diminuír a eficacia do sistema inmunitario dos individuos, orixinando un gran número de mortes.[45] Outros contaminantes como o cru, restos de plásticos e as augas residuais ameazan a vida dos mamíferos mariños.[113]
A contaminación sonora causada polas actividade antropoxénicas é outra preocupación importante para a conservación dos mamíferos mariños. Este é un problema porque a contaminación acústica baixo a auga interfire coas capacidades dalgúns mamíferos mariños de comunicarse, e localizar a predadores e presas.[114] As explosións submarinas utilízanse para diversos propósitos, como actividades militares, construcións e investigacións oceanogróficas ou xeofísicas. Poden causar danos como hemorraxias nos pulmóns, e contusións e ulceracións do tracto gastrointestinal.[92] O ruído baixo a auga é xerado por barcos, as industrias do petróleo e o gas, as investigación e uso militar do sonar e a experimentación acústica oceanográfica. Os dispositivos para o acoso acústico e aparatos de disuasión acústica usados en instalacións de acuicultura espantan os mamíferos mariños ao emitiren sons fortes e desagradables baixo a auga.[107]
Dous cambios que se están a producir na atmosfera global debido a actividades humanas ameazan os mamíferos mariños. O primeiro é o incremento de radiación ultravioleta debida á depleción da capa de ozono, o que afecta principalmente ao Antártico e outras áreas do hemisferio sur.[45] Un incremento da radicación ultravioleta ten a capacidade de diminuír a abundancia de fitoplancto, o cal forma a base da cadea trófica dos océanos.[115] O segundo efecto do cambio climático global é o quecemento global debido ao incremento dos niveis de dióxido de carbono na atmosfera. A elevación do nivel do mar, da temperatura do mar e os cambios nas correntes espérase que afecten os mamíferos mariños ao alteraren a distribución de importantes especies de presas das que se alimentan, modificando os sitios de reprodución axeitados e as rutas migratorias.[116] A cadea trófica ártica sería alterada pola case extinción ou migración dos osos polares. O xeo do océano Glacial Ártico é o hábitat do oso polar. Está diminuíndo a unha velocidade do 13% por década porque a temperatura está aumentando alí ao dobre de velocidade que no resto do mundo.[77][117] Prevese que para o ano 2050, uns dous terzos dos osos polares que hai no mundo xa terán desaparecido se o xeo mariño continúa fundindo ao ritmo actual.[118]
Protección editar

A Convención sobre a Conservación de Especies Migratorias e Animais Silvestres de 1979 (CMS) é a única organización global dedicada a conservar moitas especies animais, entre as cales se inclúen os mamíferos mariños.[119][120] Dos acordos aos que se chegou, tres deles tratan da conservación dos mamíferos mariños: ACCOBAMS, ASCOBANS e o Acordo do Waddenzee.[121] En 1982, a Convención das Nacións Unidas sobre a Lei do Mar (LOSC) adoptou un enfoque de prevención da contaminación para axudar a conservación, a cal adoptaron tamén moitas outras convencións desa época.[122]
O Acordo para a Conservación dos Cetáceos no mar Negro, mar Mediterráneo e área do Atlántico contigua (ACCOBAMS) de 1996, protexe especificamente os cetáceos na área do Meiterráneo, e "mantén un status favorable", e unha acción directa contra a caza de baleas.[122] Hai 23 estados signatarios.[123] O Acordo para a Conservación de Pequenos Cetáceos dos mares Báltico e Norte (ASCOBANS) foi adoptado xunto co ACCOBAMS para establecer unha área de protección especial para os cetáceos crecentemente ameazados de Europa.[122] Outros esforzos anti caza de baleas son a moratoria de 10 anos de 1986 do IWC sobre a caza de baleas,[124] e un acordo ambiental (un tipo de lei internacional), a Convención Internacional para a Regulación da Caza de Baleas, que controla a caza de baleas comercial, científica e de subsistencia.[125]
O Acordo sobre a Conservación de Focas no Waddenzee, que entrou en vigor en 1991, prohibe matar ou acosar as focas no Waddenzee, especialmente a poboación de Phoca vitulina.[126]
En 1973 Canadá, Dinamarca (Groenlandia), Noruega (Svalbard), os Estados Unidos e a antiga Unión Soviética asinaron o Acordo para a Conservación dos Osos Polares, que facía ilegal a caza de osos polares non regulada desde avións e quebraxeos, e protexía os sitios de migración, alimentación e hibernación.[127]
Varias organizacións non gobernamentais participan no activismo da conservación mariña, dirixindo a súa atención e axuda aos diversos problemas da conservación mariña, como a polución, caza de baleas, capturas accidentais etc. Entre as organizacións máis notables está Greenpeace, que se centrou na sobrepesca e na caza de baleas entre outras cousas, e a Sea Shepherd Conservation Society que utilizan tácticas de acción directa para poñer en evidencia as actividades ilegais.[128]
Ademais destes acordos internacionais, os diferentes países adoptaron lexislación interna protectora. Por exemplo, nos Estados Unidos están protexidos pola Lei de Protección de Mamíferos Mariños de 1972 (MMPA).[129][130]:5[130]:10. A Comisión de Mamíferos Mariños (MMC) establece e revisa as políticas e fai recomendacións para unha mellor aplicación da MMPA.[130]:7
Como alimento para os humanos editar

Durante miles de anos os pobos indíxenas do Ártico dependeron para o seu sustento da carne de balea. Hoxe, esta carne obtense da caza legal non comercial que ocorre dúas veces ao ano en primavera e outono. A carne é almacenada e comida o longo do inverno.[131] A pel e a graxa (muktuk) obtida da balea de Groenlandia, a beluga ou o narval son tamén moi valoradas, e cómense crúas ou cociñadas. A caza de baleas tamén se practicou nas illas Feroe no Atlántico Norte desde o tempo dos primeiros asentamentos noruegueses (viquingos) nas illas. Aínda se matan uns 1 000 caldeiróns anualmente, principalmente durante o verán.[132][133] Hoxe, a carne de golfiño consómese nuns poucos países do mundo, entre os que están o Xapón[134][135] e o Perú (onde se denomina chancho marino, é dicir, "porco mariño").[136] Nalgunhas partes do mundo, como Taiji, Xapón as illas Feroe, os golfiños son considerados tradicionalmente un alimento, e son cazados con arpóns ou reunidos nunha baía e matados.[134]
Hai riscos para a saúde asociados co consumo de carne de golfiños no Xapón despois de que as análises da carne de golfiño indicaron que contiña altos niveis de metilmercurio.[135][137] Porén, non se coñece ningún caso de envelenamento por mercurio como resultado do consumo de carne de golfiño, aínda que o goberno xaponés continúa facendo un control das persoas que viven en áreas onde hai un consumo alto desta carne. O goberno xaponés recomenda que os nenos e as mulleres preñadas eviten comer esta carne de forma regular.[138] Existen preocupacións similares co consumo de carne de golfiño nas illas Feroe, onde a exposición prenatal ao metilmercurio e PCBs polo consumo de carne de caldeirón tivo como resultado a aparición de casos de déficits neuropsicolóxicos en nenos.[137]
A poboación das illas Feroe foi exposta en gran medida ao metilmercurio pola carne de caldeirón contaminada, que contiña niveis moi altos duns 2 mg de metilmercurio/kg. Porén, as pobaocións das illas Feroe tamén comen cantidades significativas de peixes. O estudo duns 900 nenos feroeses mostrou que a exposición prenatal ao metilmercurio tivo como resultado déficits neuropsicolóxicos aos 7 anos de idade. (fonte: Organización Mundial da Saúde [137])
As focas Pusa hispida eran antes o principal alimento dos inuit. Aínda hoxe son un importatne recurso para o pobo nunavut[139] e son tamén cazadas e comidas en Alasca. A carne de foca é unha importatne fonte de alimento para os habitantes de pequenas comunidades costeiras.[140] A graxa de foca úsase para facer aceite de foca, que se vende como un suplemento de aceite de peixe. En 2001, o 2% do aceite de foca cru canadense era procesado e vendido en tendas de produtos para a saúde do Canadá.[141]
En catividade editar
Acuarios editar
- Cetáceos

Varias especies de golfiños mantéñense en catividade, xeralmente en parques temáticos e delfinarios. Os golfiños Tursiops son as especies de golfiños máis comúns que se teñen en delfinarios, xa que son relativamente fáciles de adestrar e teñen unha longa vida en catividade. Centos de golfiños Tursiops viven en catividade ao longo do mundo, aínda que o número exacto é difícil de determinar.[142] O "sorriso" dos golfiños fixo a estes animais atraccións moi populares, xa que é unha expresión facial de benvida en humanos; porén, o sorriso é debido á falta de músculos faciais e á subseguinte falta de expresións faciais.[143]
Organizacións como World Animal Protection e a Whale and Dolphin Conservation Society fan campañas contra a práctica de manter os cetáceos, especialmente candorcas, en catividade, onde adoitan desenvolver patoloxías, como o colapso da aleta dorsal nun 60 a 90% das candorcas macho. Os animais cativos teñen unha vida moito máis reducida, como media de só uns 20 anos. Na natureza, as femias que sobreviven á infancia viven como media 46 anos, e en raros casos chegan aos 70 a 80 anos. Os machos silvestres que sobreviven á infancia viven 31 anos como media e ata 50 a 60 anos ás veces.[144] As condicións da catividade xeralmente ten pouca semellanza co hábitat natural, e os grupos sociais de baleas en catividade son estraños aos que se encontran na natureza. A vida en catividade é tamén estresante debido á necesidade de aprenderlles a realizar trucos de circo que non son parte do comportamento das candorcas na natureza salvaxe, e ao tamaño restrinxido das piscinas onde viven. As candorcas silvestres poden viaxar ata 160 km ao día, e os críticos din que os animais son demasiado grandes e intelixentes para ser especies axeitadas para a catividade.[145] Os individuos en catividade ocasionalmente son agresivos consigo mesmo, cos seus compañeiros de tanque, ou cos humanos, o que segundo os críticos é o resultado do estrés.[146] Os golfiños adoitan ser adestrados a facer varios comportamentos antropomórficos, como dicir adeus coas aletas ou dar bicos, que raramente faría un golfiño silvestre.[147]
- Pinnípedes

O gran tamaño e natureza bulideira dos pinnípedes convertéronos en atraccións populares. Algúns recintos de exhibición teñen fondos rochosos con sitios para descansar artificiais e unha piscina, mentres que outros teñen pens con refuxios elevados pequenos rochosos onde os animais poden mergullarse en piscinas. Outros máis elaborados conteñen piscinas profundas que poden verse por debaixo do nivel da auga con sitios para descansar de cemento que imitan rochas. A especie máis común de pinnípedes mantida en catividade é o león mariño californiano (Zalophus californianus), xa que é abundante e doado de adestrar.[148] Estes animais son usados para realizar trucos e entreter os visitantes.[149] Outras especies frecuentes en catividade son a Halichoerus grypus e a Phoca vitulina. Os animais máis grandes como as morsas (Odobaenus rosmarus) e os leóns mariños de Steller (Eumetopias jubatus) son moito menos comúns.[148] Os pinnípides son populares porque foron convertidos en personaxes similares aos de Disney, e, en consecuencia, a xente a miúdo os antropomorfiza cunha curiosa e divertida natureza brincadora.[150]
Algunhas organizacións, como a Humane Society of the United States e a World Animal Protection, presentan obxeccións a manter en catividade pinnípedes e outros mamíferos mariños. Afirman que os recintos de exhibición non son dabondo grandes para estes animais que evolucionaron para ser migratorios, e unha piscina nunca pode substituír o tamaño e biodiversidade do océano. Tamén se opoñen ao uso de leóns mariños en espectáculos, porque consideran que os trucos que realizan son "variacións esaxeradas dos seus comportamentos naturais" e distraen a audiencia da contorna non natural onde está o animal.[151]
- Londras mariñas
As londras mariñas poden zafarse bastante ben en catividade, e son exhibidas nuns 40 acuarios públicos e zoos.[63] O Acuario de Seattle foi a primeira institución que conseguiu criar londras mariñas desde a concepción á idade adulta.[152] Algúns vídeos sobre londras mariñas en YouTube chegaron a ter millóns de visualizacións.[153][154][155] O público considera a miúdo que as londras mariñas teñen unha "vida familiar feliz", mais iso é unha antropomorfización.[156]
- Sirenios
O manatí máis vello en catividade foi un chamado Snooty,[157] do Parker Manatee Aquarium do Museo do sur de Florida de Bradenton, Florida. Naceu en catividade en 1948 e foi criado enteiramente en catividade,[158][159] e morreu aos 69 anos.[160] Tamén hai manatís en varios zoos europeos, como o Tierpark de Berlín,[161] o zoo de Nuremberg,[162] o ZooParc de Beauval en Francia,[163] e no Acuario de Xénova en Italia.[164] No River Safari de Singapur teñen sete manatís.[165]

Uso militar editar
Países como Rusia, Estados Unidos, Ucraína e Irán desenvolveron programas para o uso de golfiños, focas ou belugas con fins militares.[166][167]
Os golfiños foron utilizados con fins militares nos Estados Unidos desde a década de 1960 e actualmente está en funcionamento o Programa de Mamíferos Mariños da Mariña dos Estados Unidos (NMMP polas súas siglas en inglés),[168] no que se utilizan golfiños Tursiops e leóns mariños de California (Zalophus californianus) para detectar minas, protexer barcos de soldados inimigos e recuperar obxectos.[166][169] A Mariña norteamericana nunca adestrou golfiños para atacar, xa que non poderían distinguir os soldados aliados dos inimigos. Creáronse cinco equipos de mamíferos mariños, cada un dos cales se especializou nunha das tres tarefas: MK4 (golfiños), MK5 (leóns mariños), MK6 (golfiños e leóns mariños), MK7 (golfiños), e MK8 (golfiños); en todos os casos MK é unha abreviación que significa marca (mark). Os equipos de golfiños foron adestrados para detectar e marcar a posición de minas ancoradas no fondo ou flotantes na columna de auga, porque os golfiños poden utilizar as súas habilidades de ecolocalización para detectar minas. O equipo dos leóns mariños rescataba equipos de probas como minas falsas ou bombas caídas de avións que estaban xeralmente fóra do alcance de mergulladores, que terían que facer múltiples mergullos. O MK6 protexe portos e barcos de mergulladores inimigos, e estivo operativo na guerra do Golfo e na do Vietnam. Os golfiños terían que nadar detrás dos mergulladores inimigos e adherir unha boia ás súas bombonas de aire, para que flotasen ata a superficie e iso alertase a soldados da Mariña próximos. Os leóns mariños terían que golpear o inimigo e tratar de evitar os seus contraataques.[170][171]
O uso de mamíferos mariños pola Mariña estadounidense, malia estar de acordo coas políticas da Mariña, continúa encontrando oposición. A política da Mariña di que só se use o reforzo positivo no adestramento militar de golfiños, e que sexan coidados segundo os estándares aceptados para o coidado de animais. O estrés inevitable do adestramento causa controversia, xa que está fóra do estilo de vida natural do animal, especialmente nos espazos confinados onde están cando non están sendo adestrados. Hai polémicas sobre o uso de buceiras e outros inhibidores, que impiden que os golfiños capturen por si sós comida cando están adestrando, e sobre o modo de transporte dos animais ou o encontro con outros golfiños, xa que son territoriais.[170][171]
Notas editar
- ↑ Jefferson, T. A.; Leatherwood, S.; Webber, M. A. (1994). Marine Mammals of the World. Food and Agriculture Department of the United Nations. pp. 1–2. ISBN 978-92-5-103292-3. OCLC 30643250.
- ↑ 2,0 2,1 2,2 Perrin, William F.; Baker, C. Scott; Berta, Annalisa; Boness, Daryl J.; Brownell Jr., Robert L.; Domning, Daryl P.; Fordyce, R. Ewan; Srembaa, Angie; Jefferson, Thomas A.; Kinze, Carl; Mead, James G.; Oliveira, Larissa R.; Rice, Dale W.; Rosel, Patricia E.; Wang, John Y.; Yamada, Tadasu, eds. (2014). "The Society for Marine Mammalogy's Taxonomy Committee List of Species and subspecies" (PDF). Consultado o 25 de xuño de 2016.
- ↑ 3,0 3,1 Kaschner, K.; Tittensor, D. P.; Ready, J.; Gerrodette, T.; Worm, B. (2011). "Current and Future Patterns of Global Marine Mammal Biodiversity". PLoS ONE 6 (5): e19653. Bibcode:2011PLoSO...619653K. PMC 3100303. PMID 21625431. doi:10.1371/journal.pone.0019653.
- ↑ Pompa, S.; Ehrlich, P. R.; Ceballos, G. (2011-08-16). "Global distribution and conservation of marine mammals". Proceedings of the National Academy of Sciences 108 (33): 13600–13605. Bibcode:2011PNAS..10813600P. doi:10.1073/pnas.1101525108.
- ↑ 5,0 5,1 Jefferson, T. A.; Webber, M. A.; Pitman, R. L. (2009). Marine Mammals of the World A Comprehensive Guide to their Identification (1st ed.). Londres: Academic Press. pp. 7–16. ISBN 978-0-12-383853-7. OCLC 326418543.
- ↑ Uhen, M. D. (2007). "Evolution of marine mammals: Back to the sea after 300 million years". The Anatomical Record 290 (6): 514–22. PMID 17516441. doi:10.1002/ar.20545.
- ↑ 7,0 7,1 Savage, R. J. G.; Domning, Daryl P.; Thewissen, J. G. M. (1994). "Fossil Sirenia of the West Atlantic and Caribbean Region. V. the Most Primitive Known Sirenian, Prorastomus sirenoides Owen, 1855". Journal of Vertebrate Paleontology 14 (3): 427–449. JSTOR 4523580. doi:10.1080/02724634.1994.10011569.
- ↑ 8,0 8,1 Schipper, J.; Chanson, J. S.; Chiozza, F.; Cox, N. A.; Hoffmann, M.; Katariya, V.; Lamoreux, J.; Rodrigues, A. S. L.; Stuart, S. N.; Temple, H. J.; Baillie, J.; Boitani, L.; Lacher, T. E.; Mittermeier, R. A.; Smith, A. T.; Absolon, D.; Aguiar, J. M.; Amori, G.; Bakkour, N.; Baldi, R.; Berridge, R. J.; Bielby, J.; Black, P. A.; Blanc, J. J.; Brooks, T. M.; Burton, J. A.; Butynski, T. M.; Catullo, G.; Chapman, R.; et al. (2008). "The Status of the World's Land and Marine Mammals: Diversity, Threat, and Knowledge" (PDF). Science 322 (5899): 225–30. Bibcode:2008Sci...322..225S. PMID 18845749. doi:10.1126/science.1165115.
- ↑ Schipper, J.; Chanson, J. S.; Chiozza, F.; Cox, N. A. (2008). "The status of the world's land and marine mammals: diversity, threat, and knowledge". Science 322 (5899): 225–230. Bibcode:2008Sci...322..225S. PMID 18845749. doi:10.1126/science.1165115.
- ↑ Castro, Peter; Huber, Michael E. (2007). Marine Biology (7th ed.). McGraw-Hill. p. 192. ISBN 978-0-07-302819-4.
- ↑ Geisler, Jonathan H.; Uden, Mark D. (2005). "Phylogenetic Relationships of Extinct Cetartiodactyls: Results of Simultaneous Analyses of Molecular, Morphological, and Stratigraphic Data". Journal of Mammalian Evolution 12 (1–2): 145–160. doi:10.1007/s10914-005-4963-8.
- ↑ Graur, D.; Higgins, G. (1994). "Molecular evidence for the inclusion of cetaceans within the order Artiodactyla" (PDF). Molecular Biology and Evolution 11 (3): 357–364. PMID 8015431. Arquivado dende o orixinal (PDF) o 04 de marzo de 2016. Consultado o 17 de agosto de 2017.
- ↑ Agnarsson, I.; May-Collado, LJ. (2008). "The phylogeny of Cetartiodactyla: the importance of dense taxon sampling, missing data, and the remarkable promise of cytochrome b to provide reliable species-level phylogenies". Molecular Phylogenetics and Evolution 48 (3): 964–985. PMID 18590827. doi:10.1016/j.ympev.2008.05.046.
- ↑ Price, SA.; Bininda-Emonds, OR.; Gittleman, JL. (2005). "A complete phylogeny of the whales, dolphins and even-toed hoofed mammals – Cetartiodactyla". Biological reviews of the Cambridge Philosophical Society 80 (3): 445–473. PMID 16094808. doi:10.1017/s1464793105006743.
- ↑ Montgelard, C.; Catzeflis, FM.; Douzery, E. (1997). "Phylogenetic relationships of artiodactyls and cetaceans as deduced from the comparison of cytochrome b and 12S RNA mitochondrial sequences". Molecular Biology and Evolution 14 (5): 550–559. PMID 9159933. doi:10.1093/oxfordjournals.molbev.a025792.
- ↑ Spaulding, M.; O'Leary, MA.; Gatesy, J. (2009). "Relationships of Cetacea -Artiodactyla- Among Mammals: Increased Taxon Sampling Alters Interpretations of Key Fossils and Character Evolution". PLoS ONE 4 (9): e7062. Bibcode:2009PLoSO...4.7062S. PMC 2740860. PMID 19774069. doi:10.1371/journal.pone.0007062.
- ↑ Thewissen, J. G. M.; Bajpai, Sunil (2001). "Whale Origins as a Poster Child for Macroevolution" (PDF). BioScience 51 (12): 1037–1049. doi:10.1641/0006-3568(2001)051[1037:WOAAPC]2.0.CO;2.
- ↑ Domning DP (2001). "The Earliest Known Fully Quadrupedal Sirenian". Nature 413 (6856): 625–627. Bibcode:2001Natur.413..625D. PMID 11675784. doi:10.1038/35098072.
- ↑ 19,0 19,1 Prins, Herbert H. T.; Gordon, Iain J., eds. (2014). "The Biological Invasion of Sirenia into Australasia". Invasion Biology and Ecological Theory. Cambridge: Cambridge University Press. p. 123. ISBN 978-1-107-03581-2. OCLC 850909221.
- ↑ Samonds, K. E.; Zalmout, I. S.; Irwin, M. T.; Krause, D. W.; Rogers, R. R.; Raharivony, L. L. (2009). "Eotheroides lambondrano, new Middle Eocene seacow (Mammalia, Sirenia) from the Mahajanga Basin, Northwestern Madagascar". Journal of Vertebrate Paleontology 29 (4): 1233–1243. doi:10.1671/039.029.0417.
- ↑ Rybczynski, N.; Dawson, M. R.; Tedford, R. H. (2009). "A semi-aquatic Arctic mammalian carnivore from the Miocene epoch and origin of Pinnipedia". Nature 458 (7241): 1021–24. Bibcode:2009Natur.458.1021R. PMID 19396145. doi:10.1038/nature07985.
- ↑ 22,0 22,1 Arnason, U.; Gullberg, A.; Janke, A.; Kullberg, M.; Lehman, N.; Petrov, E. A.; Väinölä, R. (2006). "Pinniped phylogeny and a new hypothesis for their origin and dispersal". Molecular Phylogenetics and Evolution 41 (2): 345–354. PMID 16815048. doi:10.1016/j.ympev.2006.05.022.
- ↑ Perrin 2009, pp. 861–866.
- ↑ Love, John A. (1992). Sea Otters. Golden, Colorado: Fulcrum Publishing. pp. 4–16. ISBN 978-1-55591-123-2. OCLC 25747993.
- ↑ DeMaster, Douglas P.; Stirling, Ian (8 de maio de 1981). "Ursus Maritimus". Mammalian Species 145 (145): 1–7. JSTOR 3503828. doi:10.2307/3503828.
- ↑ Kurtén, B (1964). "The evolution of the polar bear, Ursus maritimus Phipps". Acta Zoologica Fennica 108: 1–30.
- ↑ 27,0 27,1 Lindqvist, C.; Schuster, S. C.; Sun, Y.; Talbot, S. L.; Qi, J.; Ratan, A.; Tomsho, L. P.; Kasson, L.; Zeyl, E.; Aars, J.; Miller, W.; Ingolfsson, O.; Bachmann, L.; Wiig, O. (2010). "Complete mitochondrial genome of a Pleistocene jawbone unveils the origin of polar bear". Proceedings of the National Academy of Sciences 107 (11): 5053–7. Bibcode:2010PNAS..107.5053L. PMC 2841953. PMID 20194737. doi:10.1073/pnas.0914266107.
- ↑ Waits, L. P.; Talbot, S. L.; Ward, R. H.; Shields, G. F. (2008). "Mitochondrial DNA Phylogeography of the North American Brown Bear and Implications for Conservation". Conservation Biology 12 (2): 408–417. JSTOR 2387511. doi:10.1111/j.1523-1739.1998.96351.x.
- ↑ Marris, E. (2007). "Linnaeus at 300: The species and the specious". Nature 446 (7133): 250–253. Bibcode:2007Natur.446..250M. PMID 17361153. doi:10.1038/446250a.
- ↑ Kaschner, K.; Tittensor, D. P.; Ready, J.; Gerrodette, T.; Worm, B. (2011). "Current and future patterns of global marine mammal biodiversity". PLoS ONE 6 (5): e19653. Bibcode:2011PLoSO...619653K. PMC 3100303. PMID 21625431. doi:10.1371/journal.pone.0019653.
- ↑ 31,0 31,1 31,2 31,3 31,4 31,5 31,6 31,7 Berta, A; Sumich, J. L. (1999). "Exploitation and conservation". Marine Mammals: Evolutionary Biology. San Diego: Academic Press. ISBN 978-0-12-093225-2. OCLC 42467530.
- ↑ 32,0 32,1 32,2 32,3 32,4 Riedman, M. (1990). The Pinnipeds: Seals, Sea Lions, and Walruses. Los Angeles: University of California Press. ISBN 978-0-520-06497-3. OCLC 19511610.
- ↑ Whitehead, H. (2003). Sperm Whales: Social Evolution in the Ocean. Chicago: University of Chicago Press. p. 79. ISBN 978-0-226-89518-5. OCLC 51242162.
- ↑ Marsh, H.; Eros, Carole; Hugues, Joanna; Penrose, Helen (2002). Dugong: status reports and action plans for countries and territories (PDF). International Union for Conservation of Nature. ISBN 978-92-807-2130-0. OCLC 51040880. Arquivado dende o orixinal (PDF) o 07 de outubro de 2016. Consultado o 17 de agosto de 2017.
- ↑ Silverstein, Alvin; Silverstein, Virginia; Silverstein, Robert (1995). The Sea Otter. Brookfield, Connecticut: The Millbrook Press, Inc. p. 19. ISBN 978-1-56294-418-6. OCLC 30436543.
- ↑ Kenyon, Karl W. (1975). The Sea Otter in the Eastern Pacific Ocean. New York: Dover Publications. ISBN 978-0-486-21346-0. OCLC 1504461.
- ↑ Stirling, Ian (1988). "Distribution and Abundance". Polar Bears. Ann Arbor: University of Michigan Press. ISBN 0-472-10100-5.
- ↑ Lockyer, C. J. H.; Brown, S. G. (1981). "The Migration of Whales". En Aidley, D. Animal Migration. CUP Archive. p. 111. ISBN 978-0-521-23274-6.
- ↑ 39,0 39,1 Perrin 2009, p. 360.
- ↑ Lee, Jane J. (2015). "A Gray Whale Breaks The Record For Longest Mammal Migration". National Geographic. Consultado o 23 de xaneiro de 2016.
- ↑ Deutsch, C.J.; Self-Sullivan, C. & Mignucci-Giannoni, A. (2008). "Trichechus manatus". The IUCN Red List of Threatened Species 2008: e.T22103A9356917. doi:10.2305/IUCN.UK.2008.RLTS.T22103A9356917.en. Arquivado dende o orixinal o 15 de febreiro de 2019. Consultado o 23 de decembro de 2017.
- ↑ Pfeiffer, Carl J. (1997). "Renal cellular and tissue specializations in the bottlenose dolphin (Tursiops truncatus) and beluga whale (Delphinapterus leucas)" (PDF). Aquatic Mammals 23 (2): 75–84. Arquivado dende o orixinal (PDF) o 26 de abril de 2014. Consultado o 2014-04-25.
- ↑ Lockyer, Christina (1991). "Body composition of the sperm whale, Physeter cation, with special reference to the possible functions of fat depots" (PDF). Journal of the Marine Research Institute 12 (2). ISSN 0484-9019.
- ↑ Hochachka, P.; Storey, K. (1975). "Metabolic consequences of diving in animals and man". Science 187 (4177): 613–621. Bibcode:1975Sci...187..613H. ISSN 0036-8075. PMID 163485. doi:10.1126/science.163485.
- ↑ 45,0 45,1 45,2 45,3 45,4 45,5 45,6 Whitehead, H.; Reeves, R. R.; Tyack, P. L. (2000). "Science and the conversation, protection, and management of wild cetaceans". En Mann, J.; Connor, R. C. Cetacean societies : field studies of dolphins and whales. Chicago: University of Chicago Press. ISBN 978-0-226-50340-0. OCLC 42309843.
- ↑ Cranford, T. W. (2000). "In Search of Impulse Sound Sources in Odontocetes". En Au, W. W. L.; Popper, A. N.; Fay, R. R. Hearing by Whales and Dolphins. Springer Handbook of Auditory Research. New York: Springer-Verlag. ISBN 978-1-4612-7024-9. OCLC 840278009.
- ↑ Nummela, Sirpa; Thewissen, J.G.M; Bajpai, Sunil; Hussain, Taseer; Kumar, Kishor (2007). "Sound transmission in archaic and modern whales: Anatomical adaptations for underwater hearing". The Anatomical Record 290 (6): 716–733. PMID 17516434. doi:10.1002/ar.20528.
- ↑ Reidenberg, Joy S. (2007). "Anatomical Adaptations of Aquatic Mammals". The Anatomical Record 290 (6): 507–513. OCLC 255630658. PMID 17516440. doi:10.1002/ar.20541.
- ↑ Klinowska, Margaret; Cooke, Justin (1991). Dolphins, Porpoises, and Whales of the World: the IUCN Red Data Book (PDF). Columbia University Press, NY: IUCN Publications. ISBN 978-2-88032-936-5. OCLC 24110680.
- ↑ 50,0 50,1 Perrin 2009, pp. 570–572.
- ↑ U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service. "Coastal Stock(s) of Atlantic Bottlenose Dolphin: Status Review and Management Proceedings and Recommendations from a Workshop held in Beaufort, North Carolina, 13 September 1993 – 14 September 1993" (PDF). pp. 56–57.
- ↑ Gregory K. Silber, Dagmar Fertl (1995) – Intentional beaching by bottlenose dolphins (Tursiops truncatus) in the Colorado River Delta, Mexico.
- ↑ Berta, A.; Sumich, J. L.; Kovacs, K. M. (2015). Marine Mammals: Evolutionary Biology. Londres: Academic Press. p. 430. ISBN 978-0-12-397002-2. OCLC 905649783.
- ↑ Sanders, Jon G.; Beichman, Annabel C.; Roman, Joe; Scott, Jarrod J.; Emerson, David; McCarthy, James J.; Girguis, Peter R. (2015). "Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores". Nature Communications 6: 8285. Bibcode:2015NatCo...6E8285S. PMC 4595633. PMID 26393325. doi:10.1038/ncomms9285.
- ↑ Vogle, A. W.; Lillie, Margo A.; Piscitelli, Marina A.; Goldbogen, Jeremy A.; Pyenson, Nicholas D.; Shadwick, Robert E. (2015). "Stretchy nerves are an essential component of the extreme feeding mechanism of rorqual whales". Current Biology 25 (9): 360–361. PMID 25942546. doi:10.1016/j.cub.2015.03.007.
- ↑ Goldbogen, Jeremy A. (2010). "The Ultimate Mouthful: Lunge Feeding in Rorqual Whales". American Scientist 98 (2): 124. JSTOR 27859477. doi:10.1511/2010.83.124 (inactivo 2017-01-30).
- ↑ Goldbogen, J. A.; Calambokidis, J.; Oleson, E.; Potvin, J.; Pyenson, N. D.; Schorr, G.; Shadwick, R. E. (2011). "Mechanics, hydrodynamics and energetics of blue whale lunge feeding: efficiency dependence on krill density". Journal of Experimental Biology 214 (Pt 1): 131–146. PMID 21147977. doi:10.1242/jeb.048157.
- ↑ Perrin 2009, pp. 806–813.
- ↑ 59,0 59,1 Reitherman, Bruce (Producer and photographer) (1993). Waddlers and Paddlers: A Sea Otter Story–Warm Hearts & Cold Water (Documentary). U.S.A.: PBS.
- ↑ Nickerson, p. 21
- ↑ Haley, D., ed. (1986). "Sea Otter". Marine Mammals of Eastern North Pacific and Arctic Waters (2nd ed.). Seattle, Washington: Pacific Search Press. ISBN 978-0-931397-14-1. OCLC 13760343.
- ↑ "Sea otter". BBC. Arquivado dende o orixinal o 11 de setembro de 2017. Consultado o 2007-12-31.
- ↑ 63,0 63,1 63,2 63,3 63,4 VanBlaricom, Glenn R. (2001). Sea Otters. Stillwater, MN: Voyageur Press Inc. pp. 22, 33, 69. ISBN 978-0-89658-562-1. OCLC 46393741.
- ↑ Mangel, J. C.; Whitty, T.; Medina-Vogel, G.; Alfaro-Shigueto, J.; Cáceres, C.; Godley, B. J. (2010). "Latitudinal variation in diet and patterns of human interaction in the marine otter". Marine Mammal Science 27 (2): 14–25. doi:10.1111/j.1748-7692.2010.00414.x.
- ↑ Lavinge, D. M.; Kovacs, K. M.; Bonner, W. N. (2001). "Seals and Sea lions". En MacDonald, D. The Encyclopedia of Mammals (2nd ed.). Oxford University Press. pp. 147–55. ISBN 978-0-7607-1969-5. OCLC 48048972.
- ↑ "Arctic Bears". PBS Nature. 17 de febreiro de 2008. Arquivado dende o orixinal o 16 de xuño de 2008. Consultado o 17 de agosto de 2017.
- ↑ Amstrup, Steven C.; Marcot, Bruce G.; Douglas, David C. (2007). Forecasting the range-wide status of polar bears at selected times in the 21st Century (PDF). Reston, Virginia: U.S. Geological Survey.
- ↑ 68,0 68,1 Hemstock, Annie (1999). The Polar Bear. Manakato, MN: Capstone Press. pp. 24–27. ISBN 978-0-7368-0031-0. OCLC 38862448.
- ↑ Matthews, Downs (1993). Polar Bear. San Francisco: Chronicle Books. ISBN 978-0-8118-0204-8. OCLC 488971350.
- ↑ Dyck, M. G.; Romberg, S. (2007). "Observations of a wild polar bear (Ursus maritimus) successfully fishing Arctic charr (Salvelinus alpinus) and Fourhorn sculpin (Myoxocephalus quadricornis)". Polar Biology 30 (12): 1625–1628. doi:10.1007/s00300-007-0338-3.
- ↑ Marsh, Helene; O'Shea, Thomas J.; Reynolds III, John E. (2012). Ecology and Conservation of the Sirenia: Dugongs and Manatees. Cambridge: Cambridge University Press. p. 112. ISBN 978-0-521-88828-8. OCLC 773872519.
- ↑ Marsh, Helene. "Dugongidae". Fauna of Australia 1. Canberra: Australian Government Public Service. ISBN 978-0-644-06056-1. OCLC 27492815.
- ↑ Allen, Aarin Conrad; Keith, Edward O. (2015). "Using the West Indian Manatee (Trichechus manatus) as a Mechanism for Invasive Aquatic Plant Management in Florida". Journal of Aquatic Plant Management 53: 95–104.
- ↑ Estes, J. A.; Tinker, M. T.; Williams, T. M.; Doak, D. F. (1998). "Killer Whale Predation on Sea Otters Linking Oceanic and Nearshore Ecosystems". Science 282 (5388): 473–476. Bibcode:1998Sci...282..473E. ISSN 0036-8075. PMID 9774274. doi:10.1126/science.282.5388.473.
- ↑ "Aquatic Species at Risk – Species Profile – Sea Otter". Fisheries and Oceans Canada. Consultado o 29 de novembro de 2007.
- ↑ Lepak, Jesse M.; Kraft, Clifford E., Weidel, Brian C. (marzo de 2006). "Rapid food web recovery in response to removal of an introduced apex predator" (PDF). Canadian Journal of Fisheries and Aquatic Sciences 63 (3): 569–575. ISSN 0706-652X.
- ↑ 77,0 77,1 Lunn, Nicholas J.; Servanty, Sabrina; Regehr, Eric V.; Converse, Sarah J.; Richardson, Evan; Stirling, Ian (2016). "Demography of an apex predator at the edge of its range – impacts of changing sea ice on polar bears in Hudson Bay". Ecological Applications 26 (5): 1302–1320. PMID 27755745. doi:10.1890/15-1256.
- ↑ 78,0 78,1 78,2 Stirling, Ian; Guravich, Dan (1988). Polar Bears. Ann Arbor, MI: University of Michigan Press. pp. 27–28. ISBN 978-0-472-10100-9. OCLC 757032303.
- ↑ Amstrup, Steven C.; Marcot, Bruce G.; Douglas, David C. (2007). Forecasting the range-wide status of polar bears at selected times in the 21st Century (PDF). Reston, Virginia: U.S. Geological Survey.
- ↑ Barre, Lynne M.; Norberg, J. B.; Wiles, Gary J. (2005). Conservation Plan for Southern Resident Killer Whales (Orcinus orca) (PDF). Seattle: National Marine Fisheries Service (NMFS) Northwest Regional Office. p. 18. Arquivado dende o orixinal (PDF) o 26 de xuño de 2008. Consultado o 17 de agosto de 2017.
- ↑ Pyle, Peter; Schramm, Mary Jane; Keiper, Carol; Anderson, Scot D. (1999). "Predation on a white shark (Carcharodon carcharias) by a killer whale (Orcinus orca) and a possible case of competitive displacement" (PDF). Marine Mammal Science 15 (2): 563–568. doi:10.1111/j.1748-7692.1999.tb00822.x. Arquivado dende o orixinal (PDF) o 22 de marzo de 2012. Consultado o 17 de agosto de 2017.
- ↑ Visser, Ingrid N. (2005). "First Observations of Feeding on Thresher (Alopias vulpinus) and Hammerhead (Sphyrna zygaena) Sharks by Killer Whales (Orcinus orca) Specialising on Elasmobranch Prey". Aquatic Mammals 31 (1): 83–88. doi:10.1578/AM.31.1.2005.83.
- ↑ Ford, J. K. B.; Reeves, R. R. (2008). "Fight or flight: antipredator strategies of baleen whales". Mammal Review 38 (1): 50–86. doi:10.1111/j.1365-2907.2008.00118.x.
- ↑ Heimlich, Sara; Boran, James (2001). Killer Whales. Stillwater, Minnesota: Voyageur Press. ISBN 978-0-89658-545-4. OCLC 46973039.
- ↑ Springer, A. M. (2003). "Sequential megafaunal collapse in the North Pacific Ocean: An ongoing legacy of industrial whaling?". Proceedings of the National Academy of Sciences 100 (21): 12223–12228. Bibcode:2003PNAS..10012223S. PMC 218740. PMID 14526101. doi:10.1073/pnas.1635156100.
- ↑ Demaster, D; Trites, A; Clapham, P; Mizroch, S; Wade, P; Small, R; Hoef, J (2006). "The sequential megafaunal collapse hypothesis: Testing with existing data". Progress in Oceanography 68 (2–4): 329–342. Bibcode:2006PrOce..68..329D. doi:10.1016/j.pocean.2006.02.007.
- ↑ Estes, J. A.; Doak, D. F.; Springer, A. M.; Williams, T. M. (2009). "Causes and consequences of marine mammal population declines in southwest Alaska: a food-web perspective". Philosophical Transactions of the Royal Society B: Biological Sciences 364 (1524): 1647–1658. PMC 2685424. PMID 19451116. doi:10.1098/rstb.2008.0231.
- ↑ 88,0 88,1 88,2 Roman, J.; McCarthy, J. J. (2010). Roopnarine, Peter, ed. "The Whale Pump: Marine Mammals Enhance Primary Productivity in a Coastal Basin". PLoS ONE 5 (10): e13255. Bibcode:2010PLoSO...513255R. PMC 2952594. PMID 20949007. doi:10.1371/journal.pone.0013255.
- ↑ Roman, Joe; Estes, James A.; Morissette, Lyne; Smith, Craig; Costa, Daniel; McCarthy, James; Nation, J.B.; Nicol, Stephen; Pershing, Andrew; Smetacek, Victor (2014). "Whales as marine ecosystem engineers". Frontiers in Ecology and the Environment 12 (7): 377–385. doi:10.1890/130220.
- ↑ Smith, Craig R.; Baco, Amy R. (2003). "Ecology of Whale Falls at the Deep-Sea Floor" (PDF). Oceanography and Marine Biology: an Annual Review 41: 311–354.
- ↑ Fujiwara, Yoshihiro; Kawato, Masaru; Yamamoto, Tomoko; Yamanaka, Toshiro; Sato-Okoshi, Waka; Noda, Chikayo; Tsuchida, Shinji; Komai, Tomoyuki; Cubelio, Sherine S.; Sasaki, Takenori; Jacobsen, Karen; Kubokawa, Kaoru; Fujikura, Katsunori; Maruyama, Tadashi; Furushima, Yasuo; Okoshi, Kenji; Miyake, Hiroshi; Miyazaki, Masayuki; Nogi, Yuichi; Yatabe, Akiko; Okutani, Takashi (2007). "Three-year investigations into sperm whale-fall ecosystems in Japan". Marine Ecology 28 (1): 219–230. doi:10.1111/j.1439-0485.2007.00150.x.
- ↑ 92,0 92,1 92,2 92,3 92,4 92,5 Clapham, P. J.; Young, S. B.; Brownell, R. L. (1999). "Baleen whales: Conservation issues and the status of the most endangered populations". Mammal Review 29: 37–62. doi:10.1046/j.1365-2907.1999.00035.x.
- ↑ "History of Whaling". The Húsavík Whale Museum. Arquivado dende o orixinal o 21 de xuño de 2009. Consultado o 16 de maio de 2010.
- ↑ "Modern Whaling". The Húsavík Whale Museum. Arquivado dende o orixinal o 22 de xullo de 2011. Consultado o 16 de maio de 2010.
- ↑ Baker, C. S.; Cipriano, F.; Palumbi, S. R. (1996). "Molecular genetic identification of whale and dolphin products from commercial markets in Korea and Japan". Molecular Ecology 5 (5): 671–685. doi:10.1111/j.1365-294X.1996.tb00362.x.
- ↑ Harrison, John (2008). "Fur trade". Northwest Power & Conservation Council. Arquivado dende o orixinal o 10 de febreiro de 2013. Consultado o 25 de xuño de 2016.
- ↑ Haycox, Stephen W. (2002). Alaska: An American Colony. University of Washington Press. pp. 53–58. ISBN 978-0-295-98249-6. OCLC 49225731.
- ↑ 98,0 98,1 Riedman, M. (1990). The Pinnipeds: Seals, Sea Lions, and Walruses. San Francisco: University of California Press. ISBN 978-0-520-06497-3. OCLC 19511610.
- ↑ Perrin 2009, pp. 585–588.
- ↑ Beckman D. W. (2012). Marine Environmental Biology and Conservation. Jones & Bartlett Publishers. p. 315. ISBN 978-0-7637-7350-2. OCLC 613421445.
- ↑ Johnson, W. M.; Karamanlidis, A. A.; Dendrinos, P.; de Larrinoa, P. F.; Gazo, M.; González, L. M.; Güçlüsoy, H.; Pires, R.; Schnellmann, M. "Monk Seal Fact Files". monachus-guardian.org. Consultado o 9 de setembro de 2013.
- ↑ Wiig, Ø.; Amstrup, S.; Atwood, T.; Laidre, K.; Lunn, N.; Obbard, M.; Regehr, E.; Thiemann, G. (2015). "Ursus maritimus". Lista Vermella de especies ameazadas. Versión 2015.4 (en inglés). Unión Internacional para a Conservación da Natureza. Consultado o 31 de decembro de 2016.
- ↑ "Overharvest". Polar Bears International. Arquivado dende o orixinal o 04 de novembro de 2016. Consultado o 31 de decembro de 2016.
- ↑ Perrin, W. F. (1994) "Status of species" in Randall R. Reeves and Stephen Leatherwood (eds.) Dolphins, porpoises, and whales: 1994–1998 action plan for the conservation. Gland, Switzerland: International Union for Conservation of Nature and Natural Resources
- ↑ Hall, M. A. (1998). "An ecological view of the tuna—dolphin problem: impacts and trade-offs". Reviews in Fish Biology and Fisheries 8: 1–34. doi:10.1023/A:1008854816580.
- ↑ 106,0 106,1 Anderson, Paul K. (2001). "Marine Mammals in the Next One Hundred Years: Twilight for a Pleistocene Megafauna?". Journal of Mammalogy 82 (3): 623–629. JSTOR 1383601. doi:10.1093/jmammal/82.3.623.
- ↑ 107,0 107,1 107,2 107,3 107,4 Wursig, Bernd; Gailey, Glenn A. (2002). "Marine Mammals and Aquaculture: Conflicts and Potential Resolutions". En Stickney, Robert R.; McVey, James P. Responsible marine aquaculture. Wallingford, Oxon; New York: CABI. ISBN 978-0-85199-604-2. OCLC 228169018.
- ↑ Conn, P. B.; Silber, G. K. (2013). "Vessel speed restrictions reduce risk of collision-related mortality for North Atlantic right whales". Ecosphere 4 (1): art43. doi:10.1890/ES13-00004.1.
- ↑ Constantine, R.; Brunton, D. H.; Dennis, T. (2004). "Dolphin-watching tour boats change bottlenose dolphin (Tursiops truncatus) behaviour". Biological Conservation 117 (3): 299–307. doi:10.1016/j.biocon.2003.12.009.
- ↑ Rosen, D. A.; Trites, A. W. (2000). "Pollock and the decline of Steller sea lions: Testing the junk-food hypothesis". Canadian Journal of Zoology 78 (7): 1243–1250. doi:10.1139/z00-060.
- ↑ McAlpine, D. F.; Stevick, P. T.; Murison, L. D. (1999). "Increase in Extralimital Occurrences of Ice-Breeding Seals in the Northern Gulf of Maine Region: More Seals or Fewer Fish?". Marine Mammal Science 15 (3): 906–911. doi:10.1111/j.1748-7692.1999.tb00857.x.
- ↑ Hutchins, J. (1996). "Spatial and temporal variation in the density of northern cod and a review of hypotheses for the stock's collapse" (PDF). Canadian Journal of Fisheries and Aquatic Science 53 (5): 943–962. doi:10.1139/cjfas-53-5-943.
- ↑ Baker, J. R.; Jones, A. M.; Jones, T. P.; Watson, H. C. (1981). "Otter Lutra lutra L. Mortality and marine oil pollution". Biological Conservation 20 (4): 311–321. doi:10.1016/0006-3207(81)90017-3.
- ↑ Harwood, J. (2001). "Marine Mammals and their Environment in the Twenty-First Century". Journal of Mammalogy 82 (3): 630–640. JSTOR 1383602. doi:10.1644/1545-1542(2001)082<0630:MMATEI>2.0.CO;2.
- ↑ Madronich, S.; McKenzie, R. L.; Björn, L. O.; Caldwell, M. M. (1998). "Changes in biologically active ultraviolet radiation reaching the Earth's surface". Journal of Photochemistry and Photobiology B: Biology 46: 5–19. doi:10.1016/S1011-1344(98)00182-1.
- ↑ Simmonds, M. P.; Isaac, S. J. (2007). "The impacts of climate change on marine mammals: Early signs of significant problems". Oryx 41: 19. doi:10.1017/S0030605307001524.
- ↑ Stirling, Ian; Lunn, N. J.; Iacozza, J. (setembro de 1999). "Long-term trends in the population ecology of polar bears in Western Hudson Bay in relation to climatic change" (PDF). Arctic 52 (3): 294–306. doi:10.14430/arctic935. Arquivado dende o orixinal (PDF) o 28 de setembro de 2019. Consultado o 17 de agosto de 2017.
- ↑ Amstrup, S. C.; Marcot, B. G.; Douglas, D. C. (2008). DeWeaver, Eric L.; Bitz, Cecilia M.; Tremblay, L.-Bruno, eds. "Arctic Sea Ice Decline: Observations, Projections, Mechanisms, and Implications: A Bayesian Network Modeling Approach to Forecasting the 21st Century Worldwide Status of Polar Bears" (PDF). Arctic Sea Ice Decline: Observations. Arctic Sea Ice Decline: Observations, Projections, Mechanisms, and Implications (American Geophysical Union) 180: 213–268. Bibcode:2008GMS...180..213A. ISBN 9781118666470. doi:10.1029/180GM14. Arquivado dende o orixinal (PDF) o 05 de marzo de 2016. Consultado o 17 de agosto de 2017.
- ↑ "Convention on the Conservation of Migratory Species of Wild Animals" (PDF). 1979. Consultado o 7 de setembro de 2016.
- ↑ "CMS". Convention on the Conservation of Migratory Species of Wild Animals. Consultado o 7 de setembro de 2016.
- ↑ "Agreements". Convention on the Conservation of Migratory Species of Wild Animals. Consultado o 7 de setembro de 2016.
- ↑ 122,0 122,1 122,2 Ora, Nilufer (2013). Regional Co-operation and Protection of the Marine Environment Under International Law. Leiden, Netherlands: Koninklijke Brill. pp. 131–137. ISBN 978-90-04-25085-7.
- ↑ "List of Contracting Parties and Signatories" (PDF). ACCOBAMS. 2011. Arquivado dende o orixinal (PDF) o 12 de setembro de 2019. Consultado o 7 de setembro de 2016.
- ↑ "Catch Limits and Catches Taken". International Whaling Commission. Arquivado dende o orixinal o 08 de decembro de 2016. Consultado o 28 de novembro de 2016.
- ↑ International Convention for the Regulation of Whaling (PDF). International Convention for the Regulation of Whaling. Washington, D. C. 1946. pp. 1–3. Arquivado dende o orixinal (PDF) o 07 de abril de 2014. Consultado o 28 de novembro de 2016.
- ↑ Braathen, Jonette N. (1998). International Co-operation on Fisheries and Environment. TemaNord. Copenhagen: Nordic Council of Ministers. p. 45. ISBN 978-92-893-0198-5.
- ↑ "Agreement on the Conservation of Polar Bears". Oslo, Norway: IUCN/ Polar Bear Specialist Group. Arquivado dende o orixinal o 21 de decembro de 2016. Consultado o 31 de decembro de 2016.
- ↑ "Marine Conservation Organizations". MarineBio. Consultado o 28 de novembro de 2016.
- ↑ Benson, Etienne (2010). Wired Wilderness: Technologies of Tracking and the Making of Modern Wildlife. Johns Hopkins University Press. p. 147. ISBN 978-0-8018-9710-8. OCLC 502874368.
- ↑ 130,0 130,1 130,2 Marine Mammal Protection Act of 1972, 2007. Consultado o 20 de agosto de 2016. [1]
- ↑ "Native Alaskans say oil drilling threatens way of life". BBC News. xullo de 2010. Consultado o 18 de xuño de 2016.
- ↑ Nguyen, Vi (26 de novembro de 2010). "Warning over contaminated whale meat as Faroe Islands' killing continues". The Ecologist.
- ↑ Lee, Jane J. (setembro de 2014). "Faroe Island Whaling, a 1,000-Year Tradition, Comes Under Renewed Fire". National Geographic. Consultado o 18 de xuño de 2016.
- ↑ 134,0 134,1 Matsutani, Minoru (23 de setembro de 2009). "Details on how Japan's dolphin catches work". Japan Times. p. 3. Arquivado dende o orixinal o 27 de setembro de 2009. Consultado o 17 de agosto de 2017.
- ↑ 135,0 135,1 Harnell, Boyd (2007). "Taiji officials: Dolphin meat 'toxic waste'". The Japan Times. Consultado o 24 de xuño de 2016.
- ↑ Hall, Kevin G. (2003). "Dolphin meat widely available in Peruvian stores: Despite protected status, 'sea pork' is popular fare". The Seattle Times. Consultado o 18 de xuño de 2016.[Ligazón morta]
- ↑ 137,0 137,1 137,2 World Health Organisation (2008). "Guidance for identifying populations at risk from mercury exposure" (PDF). p. 36. Consultado o 29 de agosto de 2013.
- ↑ Ministry of Health, Labour and Welfare. "平成15年6月3日に公表した「水銀を含有する魚介類等の 摂食に関する注意事項」について". Ministry of Health, Labour and Welfare (en Japanese).
- ↑ "Eskimo Art, Inuit Art, Canadian Native Artwork, Canadian Aboriginal Artwork". Inuitarteskimoart.com. Arquivado dende o orixinal o 30 de maio de 2013. Consultado o 7 de maio de 2009.
- ↑ Kets de Vries, F. R. (2014). Talking to the Shaman Within Musings on Hunting. iUniverse Inc. p. 358. ISBN 978-1-4917-3034-8. OCLC 881660311.
- ↑ "5 Forslag til tiltak" (en Norwegian). Government of Norway. Arquivado dende o orixinal o 16 de abril de 2008. Consultado o 18 de xuño de 2016.
- ↑ Rose, Naoimi; Parsons, E.C.M.; Farinato, Richard. The Case Against Marine Mammals in Captivity (PDF) (4th ed.). Humane Society of the United States. pp. 13, 42, 43, 59. Arquivado dende o orixinal (PDF) o 30 de setembro de 2018. Consultado o 17 de agosto de 2017.
- ↑ White, Thomas (2007). In Defense of Dolphins: The New Moral Frontier. Malden, MA: Blackwell Publishing. p. 17. ISBN 978-1-4051-5779-7. OCLC 122974162.
- ↑ Rose, N. A. (2011). "Killer Controversy: Why Orcas Should No Longer Be Kept in Captivity" (PDF). Humane Society International and the Humane Society of the United States. Consultado o 21 de decembro de 2014.
- ↑ "Whale Attack Renews Captive Animal Debate". CBS News. 1 de marzo de 2010. Arquivado dende o orixinal o 17 de setembro de 2011. Consultado o 6 de setembro de 2015.
- ↑ Susan Jean Armstrong. Animal Ethics Reader. ISBN 978-0-415-27589-7. OCLC 51818774.
- ↑ Curtin, Susanna; Wilkes, Keith (2007). "Swimming with captive dolphins: current debates and post-experience dissonance" (PDF). International Journal of Tourism Research 9 (2): 131–146. doi:10.1002/jtr.599.
- ↑ 148,0 148,1 Larson, S. (2001). "Seals and Sea lions". En Bell, C. E. Encyclopedia of the World's Zoos 3. pp. 1148–1150. ISBN 978-1-57958-174-9. OCLC 42213993.
- ↑ Nowak, R. M. (2003). Walker's Marine Mammals of the World. Johns Hopkins University Press. pp. 80–83. ISBN 978-0-8018-7343-0. OCLC 51087217.
- ↑ Sigvaldadóttir, Sigurrós Björg (2012). "Seals as Humans—Ideas of Anthropomorphism and Disneyfication" (PDF). Selasetur Working Paper (The Wild North) (107). Arquivado dende o orixinal (PDF) o 15 de setembro de 2016. Consultado o 17 de agosto de 2017.
- ↑ "The Case Against Marine Mammals in Captivity" (PDF). Humane Society of the United States and World Animal Protection. pp. 3, 18. Arquivado dende o orixinal (PDF) o 30 de setembro de 2018. Consultado o 30 de maio de 2012.
- ↑ "Seattle Aquarium's Youngest Sea Otter Lootas Becomes a Mom". Business Wire. 19 de abril de 2000. Arquivado dende o orixinal o 19 de xuño de 2009. Consultado o 9 de marzo de 2007.
- ↑ cynthiaholmes (19 de marzo de 2007). "Otters holding hands". YouTube. Consultado o 18 de xuño de 2016.
- ↑ Sweeney, Susan; Craig, Randall (2011). Social Media for Business: 101 Ways to Grow Your Business Without Wasting Your Time. Gulf Breeze, Florida: Maximum Press. p. 86. ISBN 978-1-931644-91-4. OCLC 656846644.
- ↑ "Vancouver sea otters a hit on YouTube". CBC News. 3 de abril de 2007. Consultado o 15 de xaneiro de 2007.
- ↑ Kruuk, Hans (2006). Otters: Ecology, Behaviour and Conservation. Oxford, New York: Oxford University Press. p. 90. ISBN 978-0-19-856586-4. OCLC 137241436.
- ↑ Aronson, Claire. "Guinness World Records names Snooty of Bradenton as 'Oldest Manatee in Captivity'". bradenton.com. Bradenton Herald. Arquivado dende o orixinal o 28 de xuño de 2015. Consultado o 26 de xuño de 2015.
- ↑ Caldwell, Alicia (outubro de 2001). "He's a captive of affection". St. Petersburg Times. Consultado o 18 de xuño de 2016.
- ↑ Pitman, Craig (xullo 2008). "A manatee milestone: Snooty turning 60". Tampa Bay Times. Arquivado dende o orixinal o 06 de xuño de 2014. Consultado o 18 de xuño de 2016.
- ↑ Meller, Katie (2017). "Snooty the famous manatee dies in 'heartbreaking accident' days after his 69th birthday". Washington Post. Consultado o 27 de xullo de 2017.
- ↑ Blaszkiewitz, B. (1995). "Die Seekuhanlage im Tierpark Berlin-Friedrichsfelde". Zoologischer Garten (en alemán) 65: 175–181.
- ↑ Mühling, P. (1985). "Zum ersten Mal: Drei Seekuhgeburten in einem Zoo. Erfolgreiche Haltung und Zucht von Rundschwanz-Seekühen (Trichechus manatus)". Tiergarten Aktuell (Nuremberg) (en alemán) 1 (1): 8–16.
- ↑ "Extraordinary Animals: Manatees". Zooparc de Beauval. Arquivado dende o orixinal o 07 de agosto de 2016. Consultado o 24 de xuño de 2016.
- ↑ Eyewitness Travel Family Guide Italy: Milan & the Northwest Italy. New York: DK Publishing. 2012. ISBN 978-1-336-12080-8. OCLC 934043451.
- ↑ "Manatees move into world's largest freshwater aquarium at River Safari". The Straits Times. Consultado o 18 de xuño de 2016.
- ↑ 166,0 166,1 Lee, Jane J. (3 de maio de 2019). "Dolphins Deployed as Undersea Agents in Iraq". National Geographic News. Consultado o 18 de maio de 2019.
Using marine mammals for military purposes isn't restricted to Russia—the U.S. Navy has had a similar program since the 1960s (O uso de mamíferos mariños con propósitos militares non está restrinxido a Rusia; a Mariña dos EUA ten un programa similar desde a década de 1960)
- ↑ "Iran buys kamikaze dolphins". BBC News. 8 March 2000.
- ↑ "U.S. Navy Marine Mammal Program". Space and Naval Warfare Systems Command. Arquivado dende o orixinal o 13 de maio de 2019. Consultado o 18 de maio de 2019.
- ↑ "A Brief History of the Navy's Marine Mammal Program". Arquivado dende o orixinal o 05 de febreiro de 2006. Consultado o 18 de maio de 2019.
- ↑ 170,0 170,1 Eglan, Jared (2015). Beasts of War: The Militarization of Animals. Lulu.com. pp. 126–128. ISBN 978-1-329-51613-7.
- ↑ 171,0 171,1 Kistler, John M. (2011). Animals in the Military: From Hannibal's Elephants to the Dolphins of the U.S. Navy. Santa Barbara, California: ABC-CLIO. pp. 313–321. ISBN 978-1-59884-346-0. OCLC 741119653.
Véxase tamén editar
Bibliografía editar
- Perrin, W. F.; Wursig, B.; Thewissen, J. G. M. (2009). Encyclopedia of Marine Mammals (2nd ed.). San Diego: Academic Press. ISBN 978-0-0809-1993-5. OCLC 316226747.
Outros artigos editar
- CEMMA. Coordinadora para o Estudo dos Mamíferos Mariños de Galiza
Ligazóns externas editar
- The Marine Mammal Center Grupo conservacionista centrado nos mamíferos mariños
- The Society for Marine Mammalogy A maior organización de expertos en mamíferos mariños do mundo.
- The MarineBio Conservation Society Sitio educativo sobre vida mariña
- National Oceanographic and Atmosphere Administration Axencia que trata das condicións no océano e clima con información sobre mamíferos mariños
- Introduction to the Desmostylia Museo de Paleontoloxíay, Universidade de California – grupos extintos de mamíferos mariños