Papiliónidos
| Papilionidae | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||
| Clasificación científica | |||||||||||||||
| |||||||||||||||
| Especie tipo | |||||||||||||||
| Papilio machaon | |||||||||||||||
| Subfamilias e xéneros | |||||||||||||||
|
Hai 31 xéneros e case 600 especies:
| |||||||||||||||
Os papiliónidos (Papilionidae) son unha ampla familia de bolboretas grandes e coloridas, que comprende máis de 550 especies. Aínda que a maioría son tropicais, os membros da familia habitan en todos os continentes excepto a Antártida. A familia inclúe as bolboretas máis grandes do mundo, do xénero Ornithoptera.[1]
Os papiliónidos teñen varias características distintivas; por exemplo, as eirugas de papiliónidos teñen un órgano repugnatorio chamado osmeterio no seu protórax. O osmeterio normalmente permanece oculto, pero cando a larva é ameazada sácao fóra a través dunha amosega transversal dorsal inflándoo cun fluído.[2]
A aparencia bifurcada das ás posteriores dos papiliónidos, que poden verse cando a bolboreta está pousada coas ás estendidas, dálles unha aparencia que lembra a cola dunha andoriña. Como nome científico Linneo elixiu Papilio para o xénero tipo, que en latín significa 'bolboreta'. Como epítetos para as especies Linneo usou os nomes de heroes gregos. A especie tipo é Papilio machaon en honor de Macaón, un dos fillos de Asclepio, mencionado na Ilíada.[3]
Distribución editar
En 2005, identificáranse 552 especies actuais desta familia, distribuídas nas rexións tropicais e temperadas.[4] Varias especies habitan a altitudes que van desde o nivel do mar a altas montañas, como no caso da maioría das especies de Parnassius. A maioría dos papiliónidos e a súa maior diversidade encóntranse nos trópicos e rexións subtropicais entre os 20°N e os 20°S,[5]: especialmente no sueste de Asia, e entre os 20°N e 40°N no leste de Asia. Só se encontran 12 especies en Europa e só unha especie, Papilio machaon se encontra ns Illas Británicas.[6] En Norteamérica hai só 40 especies, incluíndo varias especies tropicais e Parnassius.[7]
O papiiliónido que vive máis ao norte é o Parnassius arcticus siberiano, que se encontra no Círculo polar ártico no noroeste de Yakutia a altitudes de 1.500 metros sobre o nivel do mar.[8] Nos Himalaias, atopáronse varias especies como Parnassius epaphus a altitudes de 6.000 metros sobre o nivel do mar.[9]:221
Morfoloxía editar
As descricións detalladas das características morfolóxicas dos Papilionidae, como indicou Bingham (1905) son as seguintes:[10]:1,2
Ovo. "Con forma de cúpula ou algo faceteado, non tan alto coma largo, algo coriáceo, opaco." (Doherty.)Larva. Robusta, lisa cunha serie de tubérculos carnosos no dorso: ás veces cunha protuberancia carnosa elevada (a denominada caparuza ou crista) no cuarto segmento. O segundo segmento ten unha abertura transversa, pola cal a larva saca cando quere un órgano carnoso glandular bifurcado e ergueito que emite un cheiro forte, penetrante e máis ben agradable.
Pupa. Variable en forma pero a maior parte das veces curvada cara atrás. É angulada, coa cabeza truncada ou arredondada e a parte psterior do abdome é liso ou tuberculado. Está adherida pola cola, normalmente nunha posición perpendicular, e ademais asegurada por unha especie de cincha de seda arredor da parte media. En Parnassius, a pupa está situada nunha rede de seda frouxa situada entre follas.
Imago. Ás de forma extraordinariamente variable. As ás posteriores moi frecuentemente teñen unha cola, que pode ser delgada, ou ancha e espatulada, pero sempre é unha extensión do termen na vea 4. Nun xénero, Armandia, o termen da á posterior está prolongado formando colas nos ápices das veas 2 e 3 e tamén da vea 4. Ás anteriores (excepto nos xéneros anormais Parnassius e Hypermnestra) coas 12 veas (todas) presentes e ademais unha vea curta interna, a vea 1 a,[11] que invariablemente remata na marxe dorsal.
Estadios de desenvolvemento do papiliónido Papilio cresphontes
-
 Ovo
Ovo -
 Larva
Larva -
 Pupa
Pupa -
 Imago
Imago




Características distintivas editar
As características clave que diferencian os Papilionidae doutras familias de bolboretas son:[1]
- O osmeterio é un órgano eversible, carnoso e bifurcado que se encontra no segmento protorácico das eirugas.
- Venación. Nos papiliónidos, a segunda vea anal 2A esténdese ata a marxe da á e non se une coa primeira vea anal, 1A. Estas veas están fusionadas noutras familias de bolboretas e nelas 2A non chega á marxe da á.
- Os escleritos do cérvix (o colo membranoso entre a cabeza e o tórax) están fusionados por debaixo do colo, onde están ancorados os músculos que moven a cabeza.
-
 Eiruga de papiliónido do Vello Mundo evertendo o seu osmeterio para defenderse.
Eiruga de papiliónido do Vello Mundo evertendo o seu osmeterio para defenderse. -
 Larva de Papilio demodocus opoñendo un corno do osmeterio evaxinado a un dedo ofensor.
Larva de Papilio demodocus opoñendo un corno do osmeterio evaxinado a un dedo ofensor. -
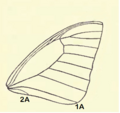 Á anterior adulta de Graphium agetes. A segunda vea anal 2A, esténdese ata a marxe da á e non se liga coa primeira vea anal, 1A.
Á anterior adulta de Graphium agetes. A segunda vea anal 2A, esténdese ata a marxe da á e non se liga coa primeira vea anal, 1A. -
 As crisálides de papiliónidos están unidas ao substrato por un cremáster e manteñen a cabeza para arriba por medio dunha faixa de seda.
As crisálides de papiliónidos están unidas ao substrato por un cremáster e manteñen a cabeza para arriba por medio dunha faixa de seda.
.jpg)



Adaptacións especiais e defensa editar
Os papiliónidos presentan mimetismo batesiano, un comportamento no cal a aparencia das bolboretas lembra moito ao de especies desagradables o que prevén que sexan depredadas. Os papiliónidos difiren de moitos animais que pesentan mimetismo. A especie Papilio glaucus mostra un polimorfismo limitado ás femias por medio de mimetismo batesiano, e outras, como Papilio canadensis non mostrran ningún tipo de mimetismo.[12]
Entre os predadores que as comen están o paxaro Agelaius phoeniceus, o escaravello Photuris pensylvanica, os lagartos Plestiodon fasciatus, as libélulas Anax junius, as arañas Misumena vatia, a mantis Tenodera sinensis, o escaravello Calosoma, e a mofeta Mephitis mephitis.[13]
Base biolóxica para os polimorfismos no mimetismo editar
Nalgunhas especies non todos os individuos son idénticos en aparencia. Por exemplo, en Papilio glaucus o ligamento ao Y determina se as femias son do tipo salvaxe (amarelas ou brancas) ou melánicas (nas que a melanina escura substitúe o fondo amarelo).[14] Esta diferenza xenética procede de que o melanismo está controlado por un só xene, que controla o nivel de dopamina no organismo. O encima BAS, que axuda á dopamina a producir a pigmentción amarela, que normalmente é a cor de fondo das ás, está suprimido. Sen a pigmentación, esta bolboreta parece principalmente negra (a forma melánica) e é un imitador batesiano do papiliónido Battus philenor. Na Papilio glaucus, que non é totalmente negra, algunhas posúen unha cor intermedia "feluxenta" e son sensibles á temperatura.[12]
Os diferentes polimorfismos (tipo salvaxe, melánico, e o intermediario 'feluxento') dependen da distribución xeográfica e da abundancia do modelo que imitan, Battus philenor, cuxa cor de ás varía dependendo da súa localiszación xeográfica.[12] Para ser confundidas con B. philenor polos predadores, a cor de fondo das ás de Papilio glaucus correspóndese coas dos B. philenor que residen na súa mesma área rexional. Diversos estudos apoian esta teoría; por exemplo, no sueste dos Estados Unidos a abundancia relativa das femias melánicas correlaciónase xeograficamente coa de B. philenor.
Mimetismo editar
Só certos grupos de papiliónidos presentan mimetismo. As especies difiren en se o mimetismo se dá nun sexo ou en ambos, e en se o mimetismo é monomórfico ou polimórfico. Un fenómeno que recibiu especial atención é o polimorfismo limitado ás femias, no cal só as femias dunha especie son miméticas e polimórficas, as cales con frecuencia imitan a bolboretas diferentes e distantemente emparentadas aposemáticas. Este polimorfismo obsérvase en Papilo dardanus, cuxa femia ten tres morfos para a patrón da cor das ás: un patrón negro e branco para o mimetismo batesiano, un patrón negro e amarelo que lembra aos machos da especie, e un patrón con manchas laranxas que lembra aos machos vellos da especie.[15] Dado que os machos da especie, que non teñen mimetismo batesiano, son depredados moito máis frecuentemente polos depredadores que as femias; unha cuestión pendente é por que as femias mostran o patrón de ás non mimético, que aparentemente diminuiría a súa fitness comparadas cos forma mimética.
Propuxéronse varias hipóteses para explicar este fenómeno, as dúas máis salientables son a hipótese da selección pseudosexual e a hipótese de evitación do macho. Na hipótese pseudosexual, as bolboretas macho aproxímanse agresivamente ás femias que parecen machos e despois moderan o seu comportamento a un comportamento sexual cando están o suficientemente preto como para identificalas como femias.[16] Na hipótese de imitación do macho, as bolboretas femias ocúltanse intentando evadirse do acoso dos machos, xa que o cortexo pode ser daniño, dura moito tempo e atrae os depredadores.[17]
Un estudo examinou a resposta dos machos ás femias de cada morfo e encontrou que os machos favorecen consistentemente aos imitadores batesianos en primeiro lugar, despois aos brancos e negros e finalmente ao morfo con manchas laranxas.[15] Os científicos concluíron que unha selección dependente da frecuencia levou a que tivesen igual éxito as tres estratexias alternativas: as femias batesianas sufrían menos depredación pero a súa fitness estaba reducida principalmente polo acoso sexual, mentres que os outros dous estaban sometidos a menor acoso sexual, pero tamén perdían fitness polos ataques dos depredadores.
Taxonomía editar
Subfamilias editar
Os xéneros dos papiliónidos existentes clasifícanse xeralmente en tres subfamilias: Baroniinae, Parnassiinae e Papilioninae; as dúas últimas divídense en tribos. Nos papiliónidos, xunto con características morfolóxicas, a elección de plantas para alimentarse e o estilo de vida ecolóxico reflicten a filoxenia e clasificación.
Baroniinae editar
Os Baroniinae son unha subfamilia monotípica, restrinxida a unha rexión moi pequena de México e son considerados a subfamilia máis basal. Baronia brevicornis é a súia única especie e considérase unha especie relicta, que comparte características cun taxon fósil, Praepapilio. Baronia é o único papiliónido que ten como planta da que se alimenta unha especie de Acacia (da familia Leguminosae).[5]
Subfamilia: Baroniinae.

Parnassiinae editar
Os Parnassiinae son unha subfamilia de bolboretas esencialmente holárticas. A gran maioría das especies, principalmente as Parnassius, poden encontrarse en hábitats montañosos. Os Parnassiinini poden tamén encontrarse noutros hábitats como os "desertos áridos (Hypermnestra), bosques húmidos (Luehdorfia) e mesmo praderías de terras baixas (Zerynthia)".[18] As tribos recoñecidas nos Parnassiinae son: Parnassiini, Zerynthiini e Luehdorfiini.
A tribo Parnassiini contén dous xéneros monotípicos, Hypermnestra, en gran medida confinada a Asia central e o xénero Parnassius (as apolos), un grupo moi distintivo con moitas especies, todas as cales son alpinas e capaces de vivir a grandes altitudes. A maioría dos Parnassius teñen dúas pequenas manchas avermelladas nas ás anteriores. O tribo Luehdorfiini contén o xénero monotípico Archon de Asia Menor e o xénero Luehdorfia da China e o Xapón. Estas dúas tribos evolucionaron cambiando as plantas das que se alimentan, mentres que a terceira tribo, Zerynthiini, mantivo a planta alimenticia arquetípica dos papiliónidos, a Aristolochia. Os Zerynthiini comprenden catro xéneros: Sericinus, Bhutanitis, Zerynthia e Allancastria.[5]:13[19]
Subfamilia: Parnassiinae.
-

Parnassius apollo,
tribo Parnassiini -

Archon apollinus,
tribo Luehdorfiini -

Zerynthia polyxena,
tribo Zerythiini -

Bhutanitis lidderdalii,
tribo Zerythiini




Papilioninae editar
As tribos recoñecidas dentro de Papilioninae son os Leptocircini, Teinopalpini, Troidini e Papilionini.
Subfamilia: Papilioninae.
-

Eurytides marcellus
tribo Leptocircini -

Teinopalpus imperialis
tribo Teinopalpini -

Trogonoptera trojana
tribo Troidini -

Papilio troilus
tribo Papilionini -

Papilio memnon
tribo Papilionini -

-

Papilio rutulus
tribo Papilionini






.jpg)
Praepapilioninae editar
Unha subfamilia adicional, Praepapilioninae, que consta do único xénero Praepapilio, comprende dúas especies de bolboretas extinguidas, cada unha das cales foi descrita por un só fósil de depósios do Eoceno medio de Colorado, Estados Unidos (Durden e Rose, 1978).[20]
Filoxenia editar
Velaquí unha filoxenia dos Papilionidae baseada en Nazari (2007):[1][19]
| Filoxenia | |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
Hoxe acéptase que subfamilia Papilioninae é monofilética.[1] Os papiliónidos da tribo Papilionini suman unhas 225 especies e fixéronse estudos sobre a coevolución coas súas plantas hóspedes e a filoxenia. Tamén se viu que as vellas clasificacións morfolóxicas eran válidas porque establecían agrupacións concordantes cos estudos. As especies que pertencen aos grupos que usan as plantas Rutaceae como hóspedes formaron dous grupos correspondentes aos taxons do Vello Mundo e as Américas. Os que se alimentan de plantas Lauraceae e Magnoliaceae forman outra agrupación, que comprende taxons de Asia e América.[21]
Os Parnassinae, igual que os Papilioninae, tamén se cre que son monofiléticos baseándose en estudos morfolóxicos pero estudos recentes baseados tanto en características morfolóxicas coma moleculares suxiren que en realidade non é así.[1] Entre os Parnassiinae, os xéneros Parnassius e Hypermnestra son extremadamente próximos baseándose en estudos moleculares[22] e son agora considerados parte da tribo Parnassiini.[19] Os dous taxóns, Archon e Luehdorfia, están moi relacionados segundo as análises de ADN nuclear e mitocondrial, e, aínda que non comparten semellanas morfolóxicas, foron agora xuntadas na tribo Luehdorfiini.[19]
A subfamilia Baroniinae está representada por unha única especie, Baronia brevicornis. Son as únicas da familia que usan plantas Fabaceae (leguminosas) como plantas hóspede das larvas. Os Baronninae e a subfamilia extinta Praepapilioninae teñen moitas semellanzas externas e son consideradas tradicionalmente as subfamilias máis primitivas e irmás do resto dos papilionidos. Investigacións recentes suxiren que isto podería non ser correcto, os Baroniinae estarían estreitamente emparentados só cos Parnassiinae, mentres que Praepapilio estaríao só cos Papilionini e ningún dos taxons sería irmán do resto dos papiliónidos.[1]
Apareamento e etapas larvarias editar
Despois do apareamento, os machos parnasinos producen unha substancia pegañosa que se usa para selar as aberturas xenitais da femia e impiden que outros machos se apareen despois.[23] Poñen un ovo no envés das follas da súa planta hóspede, repetindo o proceso moitas veces.[24] Non hai investimento parental unha vez que se puxeron os ovos.
As pupas están normalmente unidas ao substrato polo cremáster pero coa cabeza para arriba e sostida por unha faixa de seda. Porén, as apolos pupan sobre detritos no chan e tamén constrúen un casulo frouxo. Nas rexións temperadas, pasan os invernos nun estado de diapausa pupal.
Alimento editar

As eirugas de varias especies de papiliónidos aliméntanse dunha ampla variedade de plantas, e a maioría dependen de só unha das seguintes cinco familias de plantas: Aristolochiaceae, Annonaceae, Lauraceae, Umbelliferae (Apiaceae) e Rutaceae. Ao comeren algunhas destas plantas tóxicas, as eirugas secuestran ácido aristolóquico que proporciona ás eirugas e bolboretas certas cantidades deste tóxico, o que as fai tóxicas.[25] As tribos Zerynthiini (Parnassiinae), Luehdorfiini (Parnassiinae) e Troidini (Papilioninae) usan case exclusivamente plantas da familia Aristolochiaceae como hóspedes.
Os papiliónidos adultos, na súa forma de bolboreta, aliméntanse de néctar, pero tamén chuchan barro e ás veces esterco.[24]
Relación cos humanos editar
Como as bolboretas desta familia son grandes, coloridas e atractivas, foron moi procuradas polos coleccionistas de insectos. As de maior tamaño, as dos xéneros Trogonoptera, Troides e Ornithoptera, son especialmente cobizadas e son cultivadas en granxas de bolboretas para subministrar aos coleccionistas.
As larvas de moitos membros da familia aliméntanse de cítricos da familia Rutaceae. Algunhas destas atractivas bolboretas son consideradas unha praga para os hortos de cítricos.
Notas editar
- ↑ 1,0 1,1 1,2 1,3 1,4 1,5 1,6 Reed, Robert D.; Sperling, Felix A.H. (2006). "Papilionidae – The Swallowtail Butterflies". Tree of Life Web Project. Arquivado dende o orixinal o 23 de xaneiro de 2020. Consultado o 7 de novembro de 2010.
- ↑ Richards, O. W.; Davies, R.G. (1977). Imms' General Textbook of Entomology: Volume 1: Structure, Physiology and Development Volume 2: Classification and Biology. Berlin: Springer. ISBN 0-412-61390-5.
- ↑ Salmon, Michael A., Marren, Peter, Harley, Basil. The Aurelian Legacy: British Butterflies and Their Collectors. page 252. Publisher: University of California Press. 2001. ISBN 978-0-520-22963-1
- ↑ Häuser, Christoph L.; de Jong, Rienk; Lamas, Gerardo; Robbins, Robert K.; Smith, Campbell; Vane-Wright, Richard I. (28 de xullo de 2005). "Papilionidae – revised GloBIS/GART species checklist (2nd draft)". Arquivado dende o orixinal o 09 de setembro de 2010. Consultado o 8 de novembro de 2010.
- ↑ 5,0 5,1 5,2 Collins, N. Mark; Collins, Michael G. (1985). Threatened Swallowtails of the World: the IUCN red data book. IUCN Protected Area Programme Series. Gland, Switzerland and Cambridge, U.K.: IUCN. pp. 401 & 8 plates. ISBN 978-2-88032-603-6. Consultado o 22 de outubro de 2010.
- ↑ Coombs, Simon (30 de setembro de 2010). "European Butterfly checklist". butterfly-guide.co.uk. Arquivado dende o orixinal o 23 de outubro de 2010. Consultado o 11 de novembro de 2010.
- ↑ Brock, Jim P.; Kaufman, Kenn (2003). Butterflies of North America. Boston: Houghton Mifflin. ISBN 0-618-15312-8.
- ↑ Stumpe, Felix. "Parnassius arctica Eisner, 1968". Russian-Insects.com. Consultado o 9 de novembro de 2010.
- ↑ Mani, M. S. (1968). Ecology and Biogeography of High Altitude Insects. Volume 4 of Series entomologica. Springer. p. 530. ISBN 978-90-6193-114-0. Consultado o 9 de novembro de 2010.
- ↑ Bingham, C.T. (1905). The Fauna of British India including Ceylon and Burma – Butterflies (Vol 1). Londres: Taylor and Francis. p. 519. Consultado o 7 de novembro de 2010.
- ↑ A vea chámase agora 2A ou segunda vea anal nos modernos sistemas de venación.
- ↑ 12,0 12,1 12,2 Scriber, Mark; Hagen, Robert; Lederhouse, Robert (febreiro de 1996). "Genetics of Mimicry in the Tiger Swallowtail Butterflies, Papilio glaucus and P. canadensis (Lepidoptera: Papilionidae)". Evolution 50 (1): 222. JSTOR 2410795. doi:10.2307/2410795.
- ↑ Moran, Mark. "Eastern Black Swallowtail". Arquivado dende o orixinal o 01 de setembro de 2016. Consultado o 19 de outubro de 2013.
- ↑ Koch, Bernhardt; Behnecke, Bettina; ffrench-Constant, Richard H. (maio de 2000). "The molecuar basis of melanism and mimicry in a swallowtail butterfly". Current Biology 10 (10): 591–4. PMID 10837223. doi:10.1016/s0960-9822(00)00494-2.
- ↑ 15,0 15,1 Cook, S. E.; Jennifer G. Vernon; Melissa Bateson; Tim Guilford (1994). "Mate Choice in the Polymorphic African Swallowtail Butterfly, Papilio dardanus: male-like females may avoid sexual harassment" (PDF). Animal Behavior 47 (2): 389–397. doi:10.1006/anbe.1994.1053. Consultado o 15 de setembro de 2013.
- ↑ Vane-Wright, R.; C.R. Smith (1991). "Phylogenetic Relationships of Three African Swallowtail Butterflies, Papilio dardanus, P. phorcas, and P. constantinus: a cladistic analysis (Lepidoptera: Papilionidae)". Systematic Entomology. Biology of Butterflies 16 (3): 275–291. doi:10.1111/j.1365-3113.1991.tb00689.x.
- ↑ Conrad, K.F.; G Pritchard (1989). "Female Dimorphism and Physiological Colour Change in the Damselfly Argia vivida Hagen Odonata: Coenagrionidae". Canadian Journal of Zoology 67 (2): 298–304. doi:10.1139/z89-044.
- ↑ Nazari, Vazrick (2006). "Parnassius Latreille 1804". Tree of Life Web Project. Arquivado dende o orixinal o 15 de maio de 2011. Consultado o 7 de novembro de 2010.
- ↑ 19,0 19,1 19,2 19,3 19,4 Nazari, Vazrick; Sperling, Felix A.H. (2006). "Parnassiinae Duponchel, [1835]". Tree of Life. Tree of Life Web Project. Arquivado dende o orixinal o 30 de novembro de 2019. Consultado o 7 de novembro de 2010.
- ↑ Durden, C. J.; Rose, H. & Rothschild, Miriam (1978). "Butterflies from the middle Eocene: the earliest occurrence of fossil Papilionidae (Lepidoptera)". Pearce-Sellards Ser. Tex. Mem. Mus. 29 (5): 1–25..
- ↑ Aubert, J.; Legal, L; Descimon, H.; Michel, F. (1999). "Molecular phylogeny of swallowtail butterflies of the tribe Papilionini (Papilionidae, Lepidoptera)". Mol Phylogenet Evol. 12 (2): 156–167. PMID 10381318. doi:10.1006/mpev.1998.0605..
- ↑ Katoh, T.; Chichvarkhin, A.; Yagi, T.; Omoto, K. (2005). "Phylogeny and evolution of butterflies of the genus Parnassius: inferences from mitochondrial 16S and ND1 sequences". Zoolog Sci. 22 (3): 343–351. PMID 15795497. doi:10.2108/zsj.22.343..
- ↑ Ramel, Alain. "Les Papilionides, une famille en beauté". Les Insectes – Petit cours illustré d'entomologie(The Insects – A short illustrated course in Entomology). Consultado o 8 de novembro de 2010. English translation.
- ↑ 24,0 24,1 "Swallowtail Butterflies". University of Michigan. Consultado o 19 de outubro de 2013.
- ↑ von Euw, J.; Reichstein, T. & Rothschild, M. (1968). "Aristolochic acid in the swallowtail butterfly Pachlioptera aristolochiae". Isr. J. Chem. 6: 659–670. doi:10.1002/ijch.196800084..
Véxase tamén editar
| Commons ten máis contidos multimedia sobre: Papiliónidos |
| Wikispecies posúe unha páxina sobre: Papiliónidos |
Bibliografía editar
- Chattopadhyay, J. (2007). Swallowtail Butterflies, Biology & Ecology of a few Indian Species. Desh Prashan, Kolkata, India. - 134 pp. ISBN 978-81-905719-1-3.
- Glassberg, J. (2001). Butterflies through Binoculars. The West.
- Guppy, C.S. and Shepard, J.H. (2001). Butterflies of British Columbia.
- Igarashi, S. (1979). Papilionidae and their early stages [in Japanese]. 2 vols. - Tokyo, Kodansha, 218 pp., 357 pls.
- James, D.G. & Nunnallee, D. (2011). Life Histories of Cascadia Butterflies.
- Korolev V.A. (2014). Cataloges on the collection of Lepidoptera. Part II. Papilionidae. - Moscova, 387 p., 20 color tabs, ISBN 978-5-00-077163-1 [1]
- Pelham, J. (2008). Catalogue of the Butterflies of the United States and Canada.
- Pyle, R.M. (2002). The Butterflies of Cascadia.
- Rothschild, L.W. (1895). A revision of the Papilios of the Eastern Hemisphere, exclusive of Africa. Novit. Zool. 2(3):167-463 and plates pdf
- Seitz, A. (1907). 1. Gattung Papilio, Schwalbenschwänze. In: Seitz, A. (ed). Die Groß-Schmetterlinger der Erde. I. Abteilung (Die Großschmetterlinge des Palaeakrtischen Faunengebietes). 1. Band: Tagfalter. pp. 8–15. F. Lehmann, Stuttgart.
- Talbot, G. (1939). The Fauna of British India. Butterflies. Volume 1. Papilionidae and Pieridae. Taylor & Francis, Londres; xxix, 600 pp., 3 pls, 1 map.
- Tuzov V., Bogdanov P., Devyatkin A., Kaabak L., Korolev V., Murzin V., Samodurov G., Tarassov E. (1997). Guide to the butterflies of Russia and adjacent territories (Lepidoptera, Rhopalocera). Vol. 1. Sofia-Moscova: Pensoft-Press, 480 p., 79 col. plates.
- Warren, A.D., Davis, K.J., Grishin, N.V., Pelham, J.P., Stangeland, E.M. (2012). Interactive Listing of American Butterflies. [2]
Ligazóns externas editar
- "Papilionidae" en Tree of Life Arquivado 01 de marzo de 2021 en Wayback Machine.
- Familia Papilionidae en Lepidoptera.pro
- Revised GloBIS/GART Lista de especies de Papilionidae.
- GloBIS Database Arquivado 05 de novembro de 2013 en Wayback Machine. Inclúe imaxes tipo
- Páxina web do Museo de Historia Natural de Londres sobre papiliónidos afrotropicais.
- Collins, N. Mark; Morris, Michael G. (1985). Threatened Swallowtail Butterflies of the World: The IUCN Red Data Book. Gland & Cambridge: IUCN. ISBN 978-2-88032-603-6.
- Fotos e descrición de especies pouco coñecidas de pepiliónidos
- Swallowtails Papiliónidos do mundo: un resumo fotográfico.
- Bolboretas e avelaíñas de Norteamérica
- Bolboretas de Norteamérica