Pericito
Un pericito é un tipo de célula que se encontra no sistema nervioso central. Os pericitos están máis especificamente localizados arredor das capas de células endoteliais da rede de capilares do cerebro. Os pericitos exercen un papel integral no mantemento da barreira hemetocerebral e outras funcións homeostáticas e hemostáticas no cerebro.[1] Estas células son tamén un compoñente chave da unidade neurovascular, a cal inclúe as células endoteliais, astrocitos, e neuronas.[2] Os pericitos realizan funcións como a regulación do fluxo sanguíneo nos capilares, eliminación e fagocitose de refugallos celulares, e regulación da permeabilidade da barreira hematocerebral. Os pericitos poden tamén contribuír á estabilización e monitorización da maduración das células endoteliais por medio dunha comunicación directa entre a membrana celular e tamén por sinalización parácrina.[3] Recentes estudos indican tamén que a ausencia de pericitos no sistema nervioso central poden causar a rotura da barreira hematocerebral e orixinar outros cambios dexenerativos no cerebro.[1]

Morfoloxía
editar
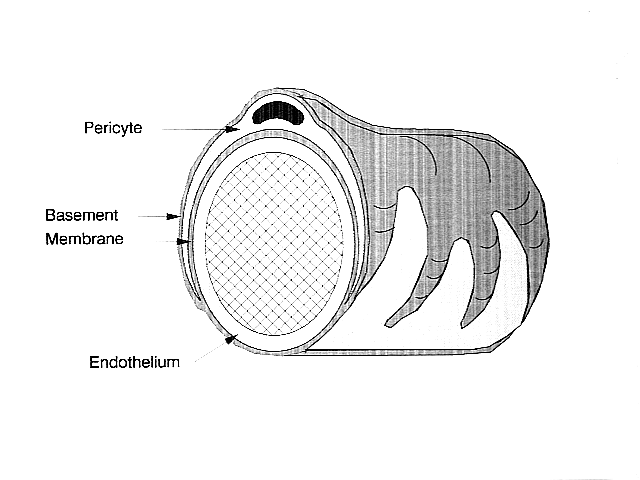
No sistema nervioso central, os pericitos envólvense arredor das células endoteliais que revisten a parte externa dos capilares. Estes dous tipos de células poden distinguirse facilmente unhas doutras polo prominente núcleo celular arredondado do pericito comparado co núcleo plano e alongado das células endoteliais.[2] Os pericitos tamén proxectan extensións con forma de dedo que rodean a parede do capilar, o que permite ás células regular o fluxo sanguíneo capilar.[1] Os pericitos e as células endoteliais comparten unha membrana basal na que se realizan moitas conexións intercelulares. Moitos tipos de moléculas de integrinas facilitan a comunicación entre os pericitos e as células endoteliais separadas pola membrana basal.[1] Os pericitos poden tamén formar conexións directas coas células veciñas nas que partes dunha célula encaixan en partes da outra, como as engrenaxes dun reloxio. Neses puntos de encaixe, poden formarse unións comunicantes, que permiten aos pericitos e ás células veciñas o intercambio de ións e outras pequenas moléculas.[1] Entre as moléculas máis importantes nestas conexións están a N-cadherina, a fibronectina, a conexina e varias integrinas.[2] Nalgunhas rexións da membrana basal, poden atoparse placas de adhesión compostas de fibronectina. Estas placas facilitan a conexión da membrana basal á estrutura do citoesqueleto composta de actina, e a membrana plasmática dos pericitos e células endoteliais.[1]
Funcións
editarBarreira hematocerebral
editarOs pericitos desempeñan un papel fundamental na formación e funcionalidade do espazo permeable selectivo entre o sistema circulatorio e o sistema nervioso central. Este espazo coñécese como barreira hematocerebral. Esta barreira está composta por células endoteliais e asegura a proteccion e funcionalidade do cerebro e sistema nervioso central. Aínda que se teorizara que os astrocitos eran cruciais para a formación postnatal desta barreira, encontrouse despois que os pericitos son en gran parte responsables desta función. Os pericitos son responsables da formación de unións herméticas e do tráfico de vesículas entre as células endoteliais. Ademais, permiten a formación da barreira ao inhibiren os efectos das células inmunitarias do sistema nervioso central (que poderían afectar á formación da barreira) e ao reduciren a expresión de moléculas que incrementan a permeabilidade vascular.[4]
Ademais da formación da barreira hematocerebral, os pericitos teñen un papel activo na súa funcionalidade ao controlaren o fluxo nos vasos sanguíneos e entre os vasos sanguíneos e o cerebro. Como células contráctiles que son, poden abrirse ou pecharse en maior ou menor grao permitindo ou non que partículas de certo tamaño flúan a través dos vasos. Esta regulación do fluxo sanguíneo é beneficiosa para a función neuronal porque impide que certas partículas do sangue entren no cerebro. Cando non están presentes os pericitos, na barreira ten lugar o proceso chamado transcitose. Isto permite que partículas de varios tamaños, entre as que están proteínas plasmáticas grandes, entren facilmente no cerebro con pouca ou ningunha regulación. Este proceso é peculiar porque a barreira hematoencefálica adoita a exercer unha forte regulación en condicións normais.[5] Deste modo, os pericitos xogan un papel fundamental para impedir que substancias químicas daniñas entren no cerebro e interrompan as funcións neurolóxicas. A funcionalidade ou disfuncionalidade dos pericitos tamén se cre que contribúe ao establecemento de enfermidades neurodexenerativas como o Alzheimer, o Parkinson e o ALS (enfermidade de Lou Gehrig). Ademais, a elasticidade do pericito é beneficiosa porque poden expandirse para reducir a inflamación e deixar que as substancias nocivas difundan fóra do cerebro.[6]
Estas células tamén son importantes no incremento da microcirculación e na redución dos efectos do envellecemento do cerebro. Nun estudo sobre ratos adultos con deficiencia de pericitos, encontrouse que a ausencia de ditas células no cerebro orixinaba danos vasculares con perda da microcirculación e do fluxo sanguíneo cerebral. Este fluxo sanguíneo é indispensable para mediar os efectos causados polo estrés, hipoxia e outras condicións que poden alterar a homeostase. Ademais, cando os pericitos non están presentes, a barreira hematoencefálica non degrada certas proteínas séricas neurotóxicas e vasculotóxicas, que así poden reforzar os cambios dexenerativos. Entre eses cambios están a inflamación e a perda de memoria e capacidade de aprendizaxe.[7]
Anxioxénese e supervivencia das células endoteliais
editarOs pericitos están tamén asociados coa capacidade das células endoteliais de diferenciarse, multiplicarse, formar ramificacións vasculares (anxioxénese), sobrevivir aos sinais apoptóticos e viaxar polo corpo. Certos pericitos, chamados pericitos microvasculares, desenvólvense arredor das paredes dos capilares e axudan ás células endoteliais nestas funcións. Os pericitos microvasculares poderían non ser células contráctiles porque carecen das isoformas de alfa-actina; que son comúns noutras células contráctiles. Estas células comunícanse coas células endoteliais por medio de unións comunicantes e á súa vez fan que as células endoteliais proliferen ou sexan inhibidas selectivamente. Se este proceso non ocorre, poden darse hiperplasia e morfoxénese vascular anormal. Estes tipos de pericitos poden tamén fagocitar proteínas exóxenas. Isto suxire que este tipo celular podería ter derivado da microglía.[8]
É tamén importante subliñar que os pericitos manteñen unha plasticidade que lles permite diferenciárense en varios tipos de células, como células musculares lisas, fibroblastos e outras células nai mesenquimais. Esta versatilidade favorece que poidan remodelar activamente os vasos sanguíneos de todo o corpo e poidan mesturarse homoxeneamente co ambiente do tecido local.[9]
Ademais de crearen e remodelaren vasos sanguíneos de modo viable, os pericitos protexen as células endoteliais da morte por apoptoe ou da causada por substancias citotóxicas. En estudos in vivo viuse que os pericitos liberan unha hormona chamada aminopeptidase pericítica N/pAPN que pode axudar a promover a anxioxénese. Cando esta hormona se mestura con células endoteliais cerebrais e con astrocitos, os pericitos agrúpanse formando estruturas que lembran a capilares. Ademais, se no experimento estaban presentes todos os elementos necesarios con excepción dos pericitos, as células endoteliais sufrían apoptose. En conclusión, os pericitos deben estar presentes para o correcto funcionamento das células endoteliais, e os astrocitos deben estar presentes para asegurar que ambas as dúas permanezan en contacto. Se non están presentes, non se produce unha anxioxénese correcta.[10] Ademais, encontrouse que os pericitos contribúen á supervivencia das células endoteliais porque secretan a proteína Bcl-w durante as interaccións celulares. A Bcl-w é unha proteína instrumental das rutas que facilitan a expresión de VEGF-A e desincentivan a apoptose.[11] Crese que o VEGF é responsable da modulación das vías de transdución de sinais da apoptose e de inhibir a activación dos encimas que inducen a apoptose. O VEGF podería utilizar dous mecanismos bioquímicos para realizar isto, que serían a fosforilación da quinase regulatoria extracelular 1 (ERK-1), que mantén a supervivencia celular, e a inhibición da proteína quinase/c-jun-NH2 quinase activada polo estrés, que tamén promove a apoptose.[12]
Patoloxías
editarDebido ao seu importante papel no mantemento e regulación da estrutrura das células endoteliais e o fluxo sanguíneo, en moitas patoloxías obsérvanse anormalidades no funcionamento dos pericitos. Poden estar presentes en exceso, o que orixina doenzas como a hipertensión e a formación de tumores, ou pode haber deficiencia deles, o que causa enfermidades dexenerativas.
Hemanxiopericitoma
editar
O hemanxiopericitoma é un raro neoplasma vascular, ou crecemento anormal, que pode ser benigno ou maligno. Na súa forma maligna, poden producirse metátases nos pulmóns, fígado, cerebro, e extremidades. Maniféstase xeralmente no fémur e parte proximal da tibia como un sarcoma óseo, e dáse normalmente en xente vella, aínda que tamén hai casos en nenos. O hemanxiopericitoma está causado por un exceso de capas de pericitos arredor de vasos sanguíneos formados incorrectamente. A diagnose deste tumaor é difícil a causa da imposibilidade de distinguir os pericitos doutros tipos de células usando o microscopio óptico. O tratamento pode implicar a extirpación cirúrxica do tumor ou radioterapia, dependendo do nivel de penetración no óso e do estado de desenvolvemento do tumor.[13]
Retinopatía diabética
editarNa retina dos pacientes de diabetes é frecuente observar perda de pericitos, e esta perda é un factor característico das fases iniciais da retinopatía diabética. Diversos estudos atoparon que os pericitos son esenciais nos diabéticos para protexer as células endoteliais dos capilares da retina. Coa perda de pericitos, fórmanse microaneurismas nos capilares. En resposta, a retina ou ben incrementa a súa permeabilidade vascular, o que fai que o ollo inche pola formación dun edema macular, ou ben que se formen novos vasos sanguíneos que permean na membrana vítrea do ollo (que separa o humor vítreo do resto do ollo). O resultado final é a redución ou perda de visión.[14] Non está claro por que os diabéticos perden pericitos; unha hipótese é que se acumulan nos pericitos sorbitol (tóxico) e produtos finais da glicación avanzada (AGE). Debido á acumulación de glicosa, a vía do poliol incrementa o seu fluxo, e acumúlanse o sorbitol intracelular e a frutosa. Isto leva a un desequilibrio osmótico, que orixina danos celulares. A presenza de grandes cantidades de glicosa tamén produce acumulación de produtos finais da glicación avanzada, que tamén danas as células. [15]
Enfermidades neurodexenerativas
editarDescubriuse que a perda de pericitos no cerebro do adulto e dos vellos orixina a interrupción dunha axeitada perfusión cerebral e do mantemento da barreira hematoencefálica, o que causa a neurodexeneración e neuroinflamación. A apoptose de pericitos no cerebro ancián pode ser o resultado de fallos na comunicación entre factores de crecemento e receptores dos pericitos. O factor de crecemento derivado de plaquetas B (PDGF-B) é liberado polas células endoteliais nos vasos sanguíneos cerebrais e despois únese aos receptores PDGFR-Beta dos pericitos, que inician a súa proliferación e migración para un correcto mantemento dos vasos sanguíneos. Cando esta sinalización se interrompe, os pericitos sofren apoptose, o que produce moitas doenzas neurodexenerativas como a enfermidade de Alzheimer e a esclerose múltiple.[16]
Investigacións actuais
editarInteraccións entre os pericitos e o endotelio
editarAs células endoteliais e os pericitos son interdependentes, de modo que cando non hai unha comunicación adecuada entre eles poden producirse diversas patoloxías nos humanos, como as mencionadas anteriormente.[17]
Hai varias vías de comunicación entre as células endoteliais e os pericitos. A primeira é a sinalización polo factor de crecemento transformante (TGF), que está mediada polas células endoteliais. É importante para a diferenciación dos pericitos.[18][19] A sinalización por anxiopoetina 1 e Tie-2 é esencial para a maduración e estabilización das células endoteliais.[20] A sinalización celular pola vía do factor de crecemento derivado de plaquetas (PDGF) que fan as células endoteliais recruta pericitos, para que os pericitos poidan migran aos vasos en crecemento. Se esta vía se bloquea, orixínase unha deficiencia de pericitos.[21] A sinalización por esfingosina-1-fosfato (S1P) tamén axuda ao recrutamento de pericitos e comunícase por medio de receptores acoplados á proteína G. O S1P sinaliza por medio de GTPases que promoven o tráfico de N-cadherina nas membranas endoteliais, o cal reforza o contacto cos pericitos.[22]
A inhibición da vía do PDGF orixina deficiencia de pericitos, e causa hiperplasia endotelial, formación de unións celulares anormais, e retinotropía diabética.[14] A falta de pericitos causa a sobrerregulación do factor de crecemento endotelial vascular (VEGF), que orixina filtracións vasculares e hemorraxia.[23] Ademais, a anxiopoetina 2 pode actuar como antagonista de Tie-2.[24] Isto desestabiliza as células endoteliais, e diminúe a interacción entre as células endoteliais e os pericitos, e pode levar á formación de tumores.[25] De xeito similar á inhibition da vía do PDGF, a anxiopoetina 2 reduce a cantidade de pericitos, causando retinopatía diabética.[26]
Cicatrización
editarDespois dunha lesión no sistema nervioso central prodúcese unha cicatrización para conservar a integridade das células circundantes. Xeralmente, os astrocitos están asociados con esta cicatrización, que se denomina cicatrización glial. Porén, existe nesta cicatrización un compoñente non glial ou estromal, e hai evidencias que apuntan a que os pericitos perivasculares xogan un importante papel axudando á formación da cicatrización.[27]
Un experimento en ratos que foran marcados xeneticamente usando o transportador de glutamato aspartato (Glast) atopou que había dous diferentes subtipos de pericitos, que se denominaron A e B. O subtipo A comprende arredor do 10% dos pericitos da medula espiñal adulta e xoga un papel crítico na cicatrización. O subtipo B comprende o resto dos pericitos e ten desmina e actina alfa do músculo liso. Unha vez que se illaron e marcaron os dous subtipos, fíxose unha incisión na parte dorsal do cerebro para ver como cicatrizaba. Tipicamente, nunha medula espiñal non danada hai dez veces máis astrocitos ca pericitos do subtipo A.[28] Porén, dúas semanas despois dos danos, o número de pericitos xa era o dobre ca o de astrocitos. O subtipo A era tres veces máis frecuente ca o subtipo B. A principal diferenza nesta abundancia está en que o subtipo A pode despegarse das paredes dos vasos sanguíneos e pode enviar pequenas proxeccións ás células veciñas e depositar proteínas da matriz extracelular.
O tecido cicatricial está moi compartimentalizado. Os pericitos forman o núcleo do tecido cicatricial, as células ependimais forman unha segunda capa arredor do núcleo, e segue outra capa formada por astrocitos.[28]
A inhibición da xeración de pericitos do subtipo A causaba un peche defectuoso das incisións na medula espiñal, o que apoia a idea de que os pericitos son importantes para a cicatrización.
Notas
editar- ↑ 1,0 1,1 1,2 1,3 1,4 1,5 Winkler, EA.; Bell, RD.; Zlokovic, BV. (2011). "Central nervous system pericytes in health and disease.". Nat Neurosci 14 (11): 1398–405. PMID 22030551. doi:10.1038/nn.2946.
- ↑ 2,0 2,1 2,2 Dore-Duffy, P.; Cleary, K. (2011). "Morphology and properties of pericytes.". Methods Mol Biol 686: 49–68. PMID 21082366. doi:10.1007/978-1-60761-938-3_2.
- ↑ Fakhrejahani, E.; Toi, M. (2012). "Tumor angiogenesis: pericytes and maturation are not to be ignored.". J Oncol 2012: 261750. PMID 22007214. doi:10.1155/2012/261750.
- ↑ Daneman, Richard et al. (2010) Pericytes are required for blood–brain barrier integrity during embryogenesis. "nature" 468, 562-566
- ↑ Investigators Reveal Key to Blood-Brain Barrier. This passage of particles can be adverse to brain function because many of such can are toxic or harmful to some extent. GEN" (2010)http://www.genengnews.com/gen-news-highlights/investigators-reveal-key-to-blood-brain-barrier/81244067/
- ↑ Pericytes regulate blood–brain barrier (2010)
- ↑ Bell, Robert D. et al. Pericytes Control Key Neurovascular Functions and Neuronal Phenotype in the Adult Brain and during Brain Aging. "Cell" 68 Vol 3 409-427 (2010) http://www.cell.com/neuron/abstract/S0896-6273(10)00824-X
- ↑ Pericyte, Astrocyte and Basal Lamina Association with the Blood Brain Barrier (BBB) http://davislab.med.arizona.edu/content/pericyte-astrocyte-and-basal-lamina-association-blood-brain-barrier-bbb Arquivado 25 de abril de 2012 en Wayback Machine.
- ↑ Gerhardt H, Betsholtz C.(2003) Endothelial-pericyte interactions in angiogenesis. "Cell Tissue Res." 314(1), 15-23 http://www.ncbi.nlm.nih.gov/pubmed/12883993
- ↑ Ramsauer, Markus et. al. (2002) Angiogenesis of the blood–brain barrier in vitro and the function of cerebral pericytes "The FASEB Journal" 10.1096/fj.01-0814fje http://www.fasebj.org/content/early/2002/08/02/fj.01-0814fje.full.pdf
- ↑ Franco, Marcela et al. (2011) Pericytes promote endothelial cell survival through induction of autocrine VEGF-A signaling and Bcl-w expression "Blood" http://bloodjournal.hematologylibrary.org/content/118/10/2906 Arquivado 13/09/2019, en Wayback Machine.
- ↑ VEGF prevents apoptosis of human microvascular endothelial cells via opposing effects on MAPK/ERK and SAPK/JNK signaling. Exp Cell Res. 1999 Mar 15;247(2):495-504. http://www.ncbi.nlm.nih.gov/pubmed/10066377
- ↑ Gellman, Harris. "Medscape: Medscape Access". Consultado o 2 de novembro de 2011.
- ↑ 14,0 14,1 Hammes, HP.; Lin, J.; Renner, O.; Shani, M.; Lundqvist, A.; Betsholtz, C.; Brownlee, M.; Deutsch, U. (2002). "Pericytes and the pathogenesis of diabetic retinopathy.". Diabetes 51 (10): 3107–12. PMID 12351455.PMID 12351455
- ↑ Ciulla, TA.; Amador, AG.; Zinman, B. (2003). "Diabetic retinopathy and diabetic macular edema: pathophysiology, screening, and novel therapies.". Diabetes Care 26 (9): 2653–64. PMID 12941734.PMID 12941734
- ↑ Bell, Robert D.; Winkler, Abhay P. Sagare; Singh, Itender; LaRue, Barb; Deane, Rashid; Zlokovic, Berislav V. (2010). "Pericytes Control Key Neurovascular Functions and Neuronal Phenotype in the Adult Brain and during Brain Aging". Neuron 68 (3): 409–27.http://www.cell.com/neuron/abstract/S0896-6273(10)00824-X
- ↑ Armulik, A.; Abramsson, A.; Betsholtz, C. (2005). "Endothelial/pericyte interactions.". Circ Res 97 (6): 512–23. PMID 16166562. doi:10.1161/01.RES.0000182903.16652.d7.
- ↑ Carvalho, RL.; Jonker, L.; Goumans, MJ.; Larsson, J.; Bouwman, P.; Karlsson, S.; Dijke, PT.; Arthur, HM.; Mummery, CL. (2004). "Defective paracrine signalling by TGFbeta in yolk sac vasculature of endoglin mutant mice: a paradigm for hereditary haemorrhagic telangiectasia.". Development 131 (24): 6237–47. PMID 15548578. doi:10.1242/dev.01529.PMID 15548578
- ↑ Hirschi, KK.; Rohovsky, SA.; D'Amore, PA. (1998). "PDGF, TGF-beta, and heterotypic cell-cell interactions mediate endothelial cell-induced recruitment of 10T1/2 cells and their differentiation to a smooth muscle fate.". J Cell Biol 141 (3): 805–14. PMID 9566978.PMID 9566978
- ↑ Thurston, G.; Suri, C.; Smith, K.; McClain, J.; Sato, TN.; Yancopoulos, GD.; McDonald, DM. (1999). "Leakage-resistant blood vessels in mice transgenically overexpressing angiopoietin-1.". Science 286 (5449): 2511–4. PMID 10617467. doi:10.1126/science.286.5449.2511.PMID 10617467
- ↑ Bjarnegård, M.; Enge, M.; Norlin, J.; Gustafsdottir, S.; Fredriksson, S.; Abramsson, A.; Takemoto, M.; Gustafsson, E.; Fässler, R. (2004). "Endothelium-specific ablation of PDGFB leads to pericyte loss and glomerular, cardiac and placental abnormalities.". Development 131 (8): 1847–57. PMID 15084468. doi:10.1242/dev.01080.
- ↑ Paik, JH.; Skoura, A.; Chae, SS.; Cowan, AE.; Han, DK.; Proia, RL.; Hla, T. (2004). "Sphingosine 1-phosphate receptor regulation of N-cadherin mediates vascular stabilization.". Genes Dev 18 (19): 2392–403. PMID 15371328. doi:10.1101/gad.1227804.
- ↑ Hellström, M.; Gerhardt, H.; Kalén, M.; Li, X.; Eriksson, U.; Wolburg, H.; Betsholtz, C. (2001). "Lack of pericytes leads to endothelial hyperplasia and abnormal vascular morphogenesis.". J Cell Biol 153 (3): 543–53. PMID 11331305.
- ↑ Maisonpierre, PC.; Suri, C.; Jones, PF.; Bartunkova, S.; Wiegand, SJ.; Radziejewski, C.; Compton, D.; McClain, J.; Aldrich, TH. (1997). "Angiopoietin-2, a natural antagonist for Tie2 that disrupts in vivo angiogenesis.". Science 277 (5322): 55–60. PMID 9204896. doi:10.1126/science.277.5322.55.
- ↑ Zhang, L.; Yang, N.; Park, JW.; Katsaros, D.; Fracchioli, S.; Cao, G.; O'Brien-Jenkins, A.; Randall, TC.; Rubin, SC. (2003). "Tumor-derived vascular endothelial growth factor up-regulates angiopoietin-2 in host endothelium and destabilizes host vasculature, supporting angiogenesis in ovarian cancer.". Cancer Res 63 (12): 3403–12. PMID 12810677.PMID 12810677
- ↑ Hammes, HP.; Lin, J.; Wagner, P.; Feng, Y.; Vom Hagen, F.; Krzizok, T.; Renner, O.; Breier, G.; Brownlee, M. (2004). "Angiopoietin-2 causes pericyte dropout in the normal retina: evidence for involvement in diabetic retinopathy.". Diabetes 53 (4): 1104–10. PMID 15047628.PMID 15047628
- ↑ Göritz, C.; Dias, DO.; Tomilin, N.; Barbacid, M.; Shupliakov, O.; Frisén, J. (2011). "A pericyte origin of spinal cord scar tissue.". Science 333 (6039): 238–42. PMID 21737741. doi:10.1126/science.1203165.
- ↑ 28,0 28,1 Barnabé-Heider, F.; Göritz, C.; Sabelström, H.; Takebayashi, H.; Pfrieger, FW.; Meletis, K.; Frisén, J. (2010). "Origin of new glial cells in intact and injured adult spinal cord.". Cell Stem Cell 7 (4): 470–82. PMID 20887953. doi:10.1016/j.stem.2010.07.014.
Véxase tamén
editarOutros artigos
editarLigazóns externas
editar- MeshName - Pericytes [1]
- Diagram at udel.edu Arquivado 21 de febreiro de 2017 en Wayback Machine.
{kind=link}
{kind=link}